Coordinated regulation of active and repressive histone methylations by a dual-specificity histone demethylase cekdm7a from caenorhabditis elegans
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT H3K9me2 and H3K27me2 are important epigenetic marks associated with transcription repression, while H3K4me3 is associated with transcription activation. It has been shown that
active and repressive histone methylations distribute in a mutually exclusive manner, but the underlying mechanism was poorly understood. Here we identified ceKDM7A, a PHD (plant
homeodomain)- and JmjC domain-containing protein, as a histone demethylase specific for H3K9me2 and H3K27me2. We further demonstrated that the PHD domain of ceKDM7A bound H3K4me3 and H3K4me3
co-localized with ceKDM7A at the genome-wide level. Disruption of the PHD domain binding to H3K4me3 reduced the demethylase activity _in vivo_, and loss of ceKDM7A reduced the expression of
its associated target genes. These results indicate that ceKDM7A is recruited to the promoter to demethylate H3K9me2 and H3K27me2 and activate gene expression through the binding of the PHD
domain to H3K4me3. Thus, our study identifies a dual-specificity histone demethylase and provides novel insights into the regulation of histone methylation. SIMILAR CONTENT BEING VIEWED BY
OTHERS MECP2 REGULATES GENE EXPRESSION THROUGH RECOGNITION OF H3K27ME3 Article Open access 19 June 2020 SMYD5 CATALYZES HISTONE H3 LYSINE 36 TRIMETHYLATION AT PROMOTERS Article Open access
09 June 2022 DREAM COMPLEX SUPPRESSES DNA METHYLATION MAINTENANCE GENES AND PRECLUDES DNA HYPERMETHYLATION Article 13 July 2020 INTRODUCTION Histone methylation is a complex
post-translational modification regulating transcription and chromatin dynamics 1, 2, 3. Methylation can occur on many arginine and lysine residues in histone proteins 4. Each lysine can
undergo three distinct states of methylation, having one (mono), two (di), or three (tri) methyl groups covalently bonded to the amine group of the lysine side chain, and arginine can be
mono-methylated or di-methylated symmetrically and asymmetrically 5. Depending on specific residues and modification states, histone methylation can either activate or repress transcription
6, 7. In general, lysine methylation at H3K9, H3K27, and H4K20 is associated with transcriptional repression, whereas methylation at H3K4, H3K36, and H3K79 is associated with transcriptional
activation. However, the mechanism by which the active and repressive marks are set up in such a coordinated manner remains poorly understood. Similar to other post-translational
modifications, histone methylation is reversible. Methylation is added by histone methyltransferases and removed by demethylases. Up to now, more than 20 histone lysine demethylases have
been identified that can remove methyl groups from histones in a sequence- and methylation state-specific manner 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22. The histone lysine
demethylases can be divided into two groups with different catalytic mechanisms. One group has two members: LSD1 and LSD2, which can only remove di- and mono-methylation from H3K4 through an
amine oxidase reaction 8, 22. All others are JmjC-domain-containing proteins that catalyze the demethylation by a hydroxylation reaction and require both iron and α-ketoglutarate as
cofactors 23. The PHD (plant homeodomain) domain, which is about 60 amino acids in length, is a C4HC3-type zinc-finger commonly found in all eukaryotes 24. The PHD domain from ING2 and NURF
were recently found to bind H3K4me3 25, 26. Interestingly, the PHD domain from a histone demethylase SMCX/Jarid1C binds H3K9me3 15, and that in BHC80, a component of the LSD1 complex,
recognizes H3K4me0 27. These studies indicate that the PHD domain from different proteins can serve as an epigenetic mark reader to interpret different epigenetic modifications. Previously
we identified KIAA1718 (KDM7A) as a dual-specificity histone demethylase for H3K9me2 and H3K27me2 that regulates neural differentiation by controlling the expression of FGF4 28. To further
understand how KDM7A binds chromatin and regulates transcription, we studied its _Caenorhabditis elegans_ ortholog. Here we demonstrate that ceKDM7A is a histone demethylase specific for
H3K9me2 and H3K27me2. Our data also demonstrate that ceKDM7A is recruited to the promoter to activate gene expression by reading H3K4me3 through its PHD domain, revealing a mechanism for the
coordinate regulation of active and repressive histone methylations. RESULTS CEKDM7A IS A HISTONE DEMETHYLASE FOR H3K9ME2 AND H3K27ME2 The _C. elegans_ protein ceKDM7A is a member of the
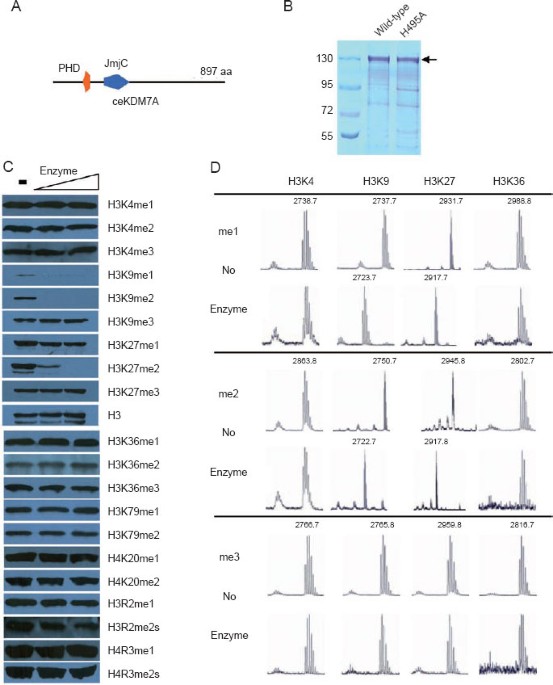
PHF2/PHF8 family, with 897 amino acids organized around two recognizable domains: PHD and JmjC (Figure 1A). To determine if ceKDM7A has histone demethylase activity _in vitro_, we expressed
and purified His-tagged full-length protein from baculovirus-infected Tn5 cells (Figure 1B). The purified protein was incubated with calf thymus histones that contain various histone
methylations. Immunoblotting demonstrated that ceKDM7A significantly reduced the signals of H3K9me1, H3K9me2, and H3K27me2 (Figure 1C). A mild reduction of H3K27me1 was also detected.
However, the levels of H3K9me3 and H3K27me3 were not affected, nor were the levels of mono-, di-, or trimethylated H3K4 and H3K36, and of methylations in other residues (Figure 1C).
MALDI-TOF mass spectrometric analysis indicated that ceKDM7A converted H3K9me1, H3K9me2, H3K27me1, and H3K27me2 to unmethylated peptides, but did not affect methylation on other residues
(Figure 1D). The demethylation by ceKDM7A requires the JmjC-domain because mutation of a conserved histidine (H495A) in the JmjC-domain abolished the activity (Figure 1B and Supplementary
information, Figure S1). These data indicate that ceKDM7A has demethylase activity for H3K9me1, H3K9me2, H3K27me1, and H3K27me2 _in vitro_. Since there are no _C. elegans_ cell lines, we
used insect cells to investigate the activity of ceKDM7A _in vivo_. Tn5 cells were infected with baculovirus expressing His-tagged full-length wild-type ceKDM7A or H495A mutant, and western
blots were probed with antibodies against a panel of histone methylations. Expression of the wild-type, but not H495A mutant, abrogated the signals of H3K9me2 and H3K27me2, but did not
affect other histone methylations examined (Figure 2A). In contrast to the _in vitro_ data, ceKDM7A expression had mild, if at all any, effect on H3K9me1 and H3K27me1. These data suggest
that ceKDM7A has demethylase activity for H3K9me2 and H3K27me2 _in vivo_, and may not have robust activity against H3K9me1 and H3K27me1 at a global scale. To determine the requirement for
the enzyme to maintain the levels of H3K9me2 and H3K27me2 in _C. elegans_, we examined the levels of these two methylations in a _C. elegans_ mutant strain tm3713. This strain harbors the
_F29B9.2_ gene knockout and does not express ceKDM7A protein (Figure 2B). We did not observe any phenotypic differences between the wild-type and mutant animals in terms of growth, life
span, and dauer formation. However, knockout of _F29B9.2_ resulted in significant elevation of H3K9me2, H3K9me1, and H3K27me2 (Figure 2C), indicating that ceKDM7A is required for proper
maintenance of these marks _in vivo_. THE PHD DOMAIN IS REQUIRED FOR DEMETHYLASE ACTIVITY AND SPECIFICALLY BINDS H3K4ME3 In addition to the JmjC domain, ceKDM7A contains a PHD domain at its
N-terminus (Figure 1A). To examine if the PHD domain is required for the demethylase activity, we infected Tn5 cells with baculovirus expressing ceKDM7A with the PHD domain deleted. While
the wild-type protein had demethylase activity against H3K9me2 and H3K27me2, the PHD-deleted mutant did not (Figure 2D). This result suggests that the PHD domain is required for the
demethylase activity _in vivo_. To determine how the PHD domain contributes to the enzymatic activity, we first examined if it binds histones, since the PHD domain was shown to be a binding
motif for both methylated and unmethylated histones 15, 25, 26. An unbiased proteomic screen demonstrated that the PHD domains from PHF2 and PHF8 specifically bind H3K4me3 29. Since the PHD
domain in ceKDM7A is highly similar to those in PHF2 and PHF8, we examined if the PHD domain in ceKDM7A has a similar binding specificity. Far western analysis indicated that the full-length
protein and the PHD domain of ceKDM7A specifically bound H3K4me3, but not other unmethylated and methylated peptides examined (Figure 3A and 3B). The specific binding was confirmed by
isothermal titration calorimetry (ITC) assay (Figure 3C). Fluorescence polarization experiments indicate that the Kd of the binding is 228 ± 11.5 μM (Figure 3D). These results indicate that
the PHD domain of ceKDM7A specifically binds H3K4me3. CO-LOCALIZATION OF H3K4ME3 AND CEKDM7A AT THE GENOME-WIDE LEVEL To study how binding of the PHD domain to H3K4me3 contributes to
enzymatic activity, we performed chromatin immunoprecipitation-coupled sequencing (ChIP-Seq) experiments. ChIP-Seq identified 695 ceKDM7A-bound genes and 698 H3K4me3-associated genes with at
least five sequence tags (Figure 4A). The numbers of the associated genes increased slightly when the cutoff of the sequence tag number was reduced to 1 (Supplementary information, Figure
S2), demonstrating high confidence of the binding. Surprisingly, there are extensive overlaps (619 genes) between ceKDM7A- and H3K4me3-associated genes (Figure 4A). Consistent with many
reported data, H3K4me3 was enriched at the transcription start site, to which ceKDM7A also bound at the genome-wide level (Figure 4B). In contrast, the levels of H3K9me2 and H3K27me2 were
low in this region. These results establish a strong correlation between ceKDM7A binding and H3K4me3 _in vivo_ and suggest that the PHD domain may bring the enzyme to its substrates by
reading H3K4me3. DISRUPTION OF THE PHD DOMAIN BINDING TO H3K4ME3 REDUCED THE DEMETHYLASE ACTIVITY IN VIVO To test if binding of the PHD domain to H3K4me3 is essential for enzymatic activity,
we mutated the six residues critical for the peptide binding as revealed by the co-crystal structure (Yang _et al._, accompanying manuscript in this issue 43). ITC experiments showed that
all mutations abolished the binding (Yang _et al._, accompanying manuscript in this issue 43), and circular dichroism spectrum indicated that the mutants maintained the secondary structure
of the enzyme (data not shown). _In vitro_ enzymatic activity assay demonstrated that the mutants had comparable enzymatic activity as that of the wild-type enzyme (Figure 5A), suggesting
that the PHD domain is not required for the enzymatic activity _per se_. However, the mutants showed decreased demethylase activity for both H3K9me2 and H3K27me2 _in vivo_ when their effect
was examined in insect cells (Figure 5B). We also made attempts to deplete H3K4me3 by its demethylase Jarid1b 30, at the same time as or prior to infection by ceKDM7A-expressing virus, but
we were unable to quantify the results when the cells were infected simultaneously or sequentially. Nevertheless, our results indicate that the PHD domain-mediated H3K4me3 binding is
required for the demethylase activity of ceKDM7A _in vivo_. LOSS OF CEKDM7A REDUCED THE EXPRESSION OF ITS ASSOCIATED GENES Since H3K9me2 and H3K27me2 are associated with transcription
repression, we examined if binding of ceKDM7A correlates with gene activation. Among 619 genes associated with ceKDM7A and H3K4me3, 142 genes with the highest levels of ceKDM7A binding were
examined by quantitative RT-PCR to compare their expression between the wild-type and _F29B9.2_ knockout worms. Among 100 genes with expression changes of more than 1.2-fold, the majority
(73 genes) were down-regulated in knockout worms, consistent with the role of ceKDM7A as a demethylase for repressive methylations (Figure 6A and Supplementary information, Table S1). When
the ChIP-Seq data of 73 down-regulated genes were re-analyzed, we observed that the levels of both H3K9me2 and H3K27me2 were reduced in ceKDM7A-bound regions, where H3K4me3 was enriched
(Figure 6B). These results suggest a hypothesis that ceKDM7A is recruited to the promoter to demethylate H3K9me2 and H3K27me2 and activate gene expression by binding to H3K4me3. DISCUSSION A
large body of evidence indicates that active and repressive histone methylations distribute in a mutually exclusive manner and that this distribution pattern is important in biology 31, 32.
Through the study of a PHD- and JmjC domain-containing protein ceKDM7A, we demonstrated that (1) ceKDM7A is a H3K9me2 and H3K27me2 histone demethylase; (2) the PHD domain binds to H3K4me3;
(3) the enzyme co-localizes with H3K4me3 in the promoter; (4) binding of H3K4me3 is required for demethylation activity _in vivo_; (5) loss of ceKDM7A reduces the expression of the
associated genes; and (6) both H3K9me2 and H3K27me2 are reduced in ceKDM7A-bound regions. Since H3K4me3 positively correlates with gene activation, and H3K9me2 and H3K27me2 are associated
with transcription repression, these results lead to a model for the coordinated regulation of active and repressive histone methylations. The model proposes a scheme of PHD domain binding
to an active histone methylation mark (H3K4me3) in the promoter, removal of repressive marks (H3K9me2 and H3K27me2) by the JmjC domain, and activation of gene expression. This model is
consistent with a recent genome-wide ChIP-Seq analysis, which indicates that H3K4 methylation primes the chromatin for gene activation 33. Coordinated regulation is a widespread phenomenon
regulating histone modifications. It can occur between methylation and acetylation 34, 35, 36, methylation and ubiquitination 37, 38, acetylation and phosphorylation 39, and arginine
methylation and lysine methylation 40. In these studies, coordinated regulation is generally mediated by protein complexes. For example, in the NuA3 HAT complex, the PHD finger of Yng1 binds
H3K4me3, which leads to H3K14 hyperacetylation by NuA3 in the same complex, and results in transcription activation of NuA3 target genes 36. Our study provides evidence that coordinated
regulation can be mediated by a single molecule. Since most of the histone-modifying enzymes carry DNA or histone-binding modules in addition to the catalytic function, the coordinated
regulation by a single molecule may apply to other epigenetic regulators. A single molecule conducting two or more related sequential tasks may have advantages over protein complexes in that
the coordinated regulation can be more efficient and more controllable, since the coordination is an intra-molecular function. MATERIALS AND METHODS REAGENTS Sources of the antibodies are
as follows: H3 monomethyl-K4 (Abcam 8895), H3 dimethy-K4 (Upstate 07-030), H3 trimethyl-K4 (Upstate 05-745), H3 monomethyl-K9 (Abcam 9045), H3 dimethyl-K9 (Upstate 07-441 and Abcam1220), H3
trimethyl-K9 (Abcam 8898), H3 monomethyl-K27 (Upstate 07-448), H3 dimethyl-K27 (Upstate 07-452 and Abcam 24684), H3 trimethyl-K27 (Upstate 07-449), H3 monomethyl-K36 (Abcam 9048), H3
dimethyl-K36 (Upstate 07-369), H3 trimethyl-K36 (Abcam 9050), H3 monomethyl-K79 (Abcam 2886), H3 dimethyl-K79 (Abcam 3594), H4 monomethyl-K20 (Abcam 9051), H4 dimethyl-K20 (Abcam 9052), H3
monomethyl-R2 (ab15584), H3 dimethyl-R2 (07-585), H4 monomethyl-R3 (ab17339-100), H4 dimethyl-R3 (ab5823-50), and H3 (abcam1791). Peptides are monomethyl-histone H3K4 (Upstate 12-563),
dimethyl-histone H3K4 (Upstate 12-460), trimethyl-histone H3K4 (Upstate 12-564), monomethyl-histone H3K9 (Upstate 12-569), dimethyl-histone H3K9 (Upstate 12-430), trimethyl-histone H3K9
(Upstate 12-568), monomethyl-histone H3K27 (Upstate 12-567), dimethyl-histone H3K27 (Upstate 12-566), trimethyl-histone H3K27 (Upstate 12-565), monomethyl-histone H3K36 (Upstate 12-570) and
the dimethyl- and trimethyl-histone H3K36 peptides are gifts from Shi Yang. The chemicals are α-ketoglutaric acid disodium salt dehydrate (Sigma Cat# 75892), ascorbic acid (Sigma Cat#
A2218), ammonium iron (II) sulfate hexahydrate (Sigma Cat# F1543), and Ni-NTA agarose (Qiagen Cat# 30210). CLONING PROCEDURES The open-reading frame of ceKDM7A was PCR amplified from Bristol
N2 cDNA and cloned into pcDNA3.1/myc-his vector (Invitrogen, Carlsbad, CA). The point mutations and deletion mutant generated by PCR were transferred into modified pFastBac1-his vector
(Invitrogen), which contains a C-terminal 6×His for affinity purification. RECOMBINANT CEKDM7A AND MUTANTS Recombinant baculovirus were generated by the Bac-to-Bac baculovirus-expressing
system (Invitrogen). HighFive (Tn5) cells were infected with baculovirus, collected 72 h later, incubated in 20 mM Hepes/NaOH (pH 7.9), 500 mM NaCl, 0.1% Triton X-100, 20% glycerol, and 1 mM
PMSF on ice for 30 min and sonicated. After centrifugation, supernatant was loaded to the Ni-NTA column and washed. The recombinant proteins were eluted and determined using SDS-PAGE
followed by Coomassie blue staining. IN VITRO DEMETHYLATION ASSAY In all, 5 μg of bulk histones or 0.2 μg of synthetic peptides was incubated with the recombinant protein in demethylation
buffer (20 mM Tris-HCl (pH 7.5), 150 mM NaCl, 50 μM (NH4)2Fe(SO4)2, 1 mM α-ketoglutarate and 2 mM ascorbic acid) for 3 h at 37 °C. Reaction mixtures were analyzed by either western blotting
using specific antibodies or mass spectrometry. GENERATION OF ANTIBODIES TO CEKDM7A Polyclonal antibodies were generated by immunizing rabbits with affinity-purified full-length ceKDM7A
(amino acids 1–897). The antibodies were affinity purified using a resin coupled with ceKDM7A. Antibody specificity was confirmed by immunoblotting and immunoprecipitation. FAR-WESTERN BLOT
ASSAY Two-branched histone H3K4, H3K9, H3K27, and H3K36 un-, mono-, di-, and tri-methylated peptides were used for far western blotting as described previously 41. Briefly, 2 μg of
two-branched peptides was loaded onto 0.1 μm pore size Protran Nitrocellulose Membranes (Whatman), dried, and stained by Ponceau as loading control. After blocking, the membranes were
incubated with 1.0 μg/ml recombinant protein overnight at 4 °C, and western blotting was performed. C. ELEGANS STRAINS The Bristol strain (N2) obtained from CGC centre was used as the
wild-type strain. tm3713 was obtained from the National BioResource Project for _C. elegans_ (Japan) and backcrossed with N2 four times. Maintenance, culturing, and genetic manipulations of
_C. elegans_ strains were carried out according to standard procedures 42 and conducted at 20 °C. CHROMATIN IMMUNOPRECIPITATION (CHIP) AND CHIP-SEQ ANALYSIS Worms were partially lyzed by
dounce, cross-linking was performed in 1% formaldehyde, and sonication was by 80 Hz, 10 s on 30 s off, 60 cycles (Sonics Vibra Cell) to shear DNA to an average fragment size of 200 to 400
bp. FA lysis buffer was used for ceKDM7A IP in the presence of yeast tRNA and BSA. After de-crosslinking and protein digestion, DNA was precipitated and quantitative PCR was performed. For
ChIP-Seq, ChIPed DNA was attached to the adaptor, amplified for 18 cycles, and subjected to sequencing by Solexa 1G Genome Analyzer. Sequence tags of mostly 35 bp were obtained and analyzed
using software maq version 0.7.1 to map ChIP-seq reads to _C. elegans_ genome (W206) and only uniquely matched reads were retained. The binding enrichment area was identified by using
software FindPeaks version 4.0.6 and annotated. The TSS (txStart) was defined according to UCSC transcript information, and 3 kb upstream and downstream of the TSS (txStart) with 200-bp
intervals. All tags were normalized by the total number of bases in the windows. RT-PCR ANALYSIS Total RNAs were extracted from worms using Trizol reagent (Invitrogen). ISOTHERMAL TITRATION
CALORIMETRY To obtain the binding affinity between F29B and histone tail modifications, purified F29B or mutants with 100 μM were titrated against various peptides (H3K4me3, H3K4me2,
H3K9me3, H3K9me2, H3K27me3 and H3K27me2) with 1 mM using VP-ITC microcalorimeter (MicroCal) at 10 °C. All proteins and peptides were prepared in a buffer containing 10 mM HEPES, pH 8.0, and
0.1 M NaCl. The data were fitted by using the software Origin 7.0. REFERENCES * Strahl BD, Allis CD . The language of covalent histone modifications. _Nature_ 2000; 403:41–45. Article CAS
PubMed Google Scholar * Bhaumik SR, Smith E, Shilatifard A . Covalent modifications of histones during development and disease pathogenesis. _Nat Struct Mol Biol_ 2007; 14:1008–1016.
Article CAS PubMed Google Scholar * Kouzarides T . Chromatin modifications and their function. _Cell_ 2007; 128:693–705. Article CAS PubMed Google Scholar * Bannister AJ, Kouzarides
T . Reversing histone methylation. _Nature_ 2005; 436:1103–1106. Article CAS PubMed Google Scholar * Bedford MT, Richard S . Arginine methylation an emerging regulator of protein
function. _Mol Cell_ 2005; 18:263–272. Article CAS PubMed Google Scholar * Li B, Carey M, Workman JL . The role of chromatin during transcription. _Cell_ 2007; 128:707–719. Article CAS
PubMed Google Scholar * Shilatifard A . Chromatin modifications by methylation and ubiquitination: implications in the regulation of gene expression. _Annu Rev Biochem_ 2006; 75:243–269.
Article CAS PubMed Google Scholar * Shi Y, Lan F, Matson C, _et al_. Histone demethylation mediated by the nuclear amine oxidase homolog LSD1. _Cell_ 2004; 119:941–953. Article CAS
PubMed Google Scholar * Lee MG, Villa R, Trojer P, _et al_. Demethylation of H3K27 regulates polycomb recruitment and H2A ubiquitination. _Science_ 2007; 318:447–450. Article CAS PubMed
Google Scholar * Lee MG, Norman J, Shilatifard A, Shiekhattar R . Physical and functional association of a trimethyl H3K4 demethylase and Ring6a/MBLR, a polycomb-like protein. _Cell_
2007; 128:877–887. Article CAS PubMed Google Scholar * Xiang Y, Zhu Z, Han G, Lin H, Xu L, Chen CD . JMJD3 is a histone H3K27 demethylase. _Cell Res_ 2007; 17:850–857. Article CAS
PubMed Google Scholar * Lan F, Bayliss PE, Rinn JL, _et al_. A histone H3 lysine 27 demethylase regulates animal posterior development. _Nature_ 2007; 449:689–694. Article CAS PubMed
Google Scholar * Christensen J, Agger K, Cloos PA, _et al_. RBP2 belongs to a family of demethylases, specific for tri-and dimethylated lysine 4 on histone 3. _Cell_ 2007; 128:1063–1076.
Article CAS PubMed Google Scholar * Cloos PA, Christensen J, Agger K, _et al_. The putative oncogene GASC1 demethylates tri- and dimethylated lysine 9 on histone H3. _Nature_ 2006;
442:307–311. Article CAS PubMed Google Scholar * Iwase S, Lan F, Bayliss P, _et al_. The X-linked mental retardation gene SMCX/JARID1C defines a family of histone H3 lysine 4
demethylases. _Cell_ 2007; 128:1077–1088. Article CAS PubMed Google Scholar * Klose RJ, Yamane K, Bae Y, _et al_. The transcriptional repressor JHDM3A demethylates trimethyl histone H3
lysine 9 and lysine 36. _Nature_ 2006; 442:312–316. Article CAS PubMed Google Scholar * Klose RJ, Yan Q, Tothova Z, _et al_. The retinoblastoma binding protein RBP2 is an H3K4
demethylase. _Cell_ 2007; 128:889–900. Article CAS PubMed Google Scholar * Whetstine JR, Nottke A, Lan F, _et al_. Reversal of histone lysine trimethylation by the JMJD2 family of
histone demethylases. _Cell_ 2006; 125:467–481. Article CAS PubMed Google Scholar * Yamane K, Tateishi K, Klose RJ, _et al_. PLU-1 is an H3K4 demethylase involved in transcriptional
repression and breast cancer cell proliferation. _Mol Cell_ 2007; 25:801–812. Article CAS PubMed Google Scholar * Yamane K, Toumazou C, Tsukada Y, _et al_. JHDM2A, a JmjC-containing H3K9
demethylase, facilitates transcription activation by androgen receptor. _Cell_ 2006; 125:483–495. Article CAS PubMed Google Scholar * Agger K, Cloos PA, Christensen J, _et al_. UTX and
JMJD3 are histone H3K27 demethylases involved in HOX gene regulation and development. _Nature_ 2007; 449:731–734. Article CAS PubMed Google Scholar * Karytinos A, Forneris F, Profumo A,
_et al_. A novel mammalian flavin-dependent histone demethylase. _J Biol Chem_ 2009; 284:17775–17782. Article CAS PubMed PubMed Central Google Scholar * Tsukada Y, Fang J,
Erdjument-Bromage H, _et al_. Histone demethylation by a family of JmjC domain-containing proteins. _Nature_ 2006; 439:811–816. Article CAS PubMed Google Scholar * Zhang Y . It takes a
PHD to interpret histone methylation. _Nat Struct Mol Biol_ 2006; 13:572–574. Article CAS PubMed Google Scholar * Wysocka J, Swigut T, Xiao H, _et al_. A PHD finger of NURF couples
histone H3 lysine 4 trimethylation with chromatin remodelling. _Nature_ 2006; 442:86–90. Article CAS PubMed Google Scholar * Shi X, Hong T, Walter KL, _et al_. ING2 PHD domain links
histone H3 lysine 4 methylation to active gene repression. _Nature_ 2006; 442:96–99. Article CAS PubMed PubMed Central Google Scholar * Lan F, Collins RE, De Cegli R, _et al_.
Recognition of unmethylated histone H3 lysine 4 links BHC80 to LSD1-mediated gene repression. _Nature_ 2007; 448:718–722. Article CAS PubMed PubMed Central Google Scholar * Huang C,
Xiang Y, Wang Y, _et al_. Dual-specificity histone demethylase KIAA1718 (KDM7A) regulates neural differentiation through FGF4. _Cell Res_ 2010; 20:154–165. Article CAS PubMed Google
Scholar * Chan DW, Wang Y, Wu M, Wong J, Qin J, Zhao Y . Unbiased proteomic screen for binding proteins to modified lysines on histone H3. _Proteomics_ 2009; 9:2343–2354. Article CAS
PubMed PubMed Central Google Scholar * Xiang Y, Zhu Z, Han G, _et al_. JARID1B is a histone H3 lysine 4 demethylase up-regulated in prostate cancer. _Proc Natl Acad Sci USA_ 2007;
104:19226–19231. Article CAS PubMed PubMed Central Google Scholar * Bernstein BE, Kamal M, Lindblad-Toh K, _et al_. Genomic maps and comparative analysis of histone modifications in
human and mouse. _Cell_ 2005; 120:169–181. Article CAS PubMed Google Scholar * Barski A, Cuddapah S, Cui K, _et al_. High-resolution profiling of histone methylations in the human
genome. _Cell_ 2007; 129:823–837. Article CAS PubMed Google Scholar * Wang Z, Zang C, Cui K, _et al_. Genome-wide mapping of HATs and HDACs reveals distinct functions in active and
inactive genes. _Cell_ 2009; 138:1019–1031. Article CAS PubMed PubMed Central Google Scholar * Dou Y, Milne TA, Tackett AJ, _et al_. Physical association and coordinate function of the
H3 K4 methyltransferase MLL1 and the H4 K16 acetyltransferase MOF. _Cell_ 2005; 121:873–885. Article CAS PubMed Google Scholar * Pray-Grant MG, Daniel JA, Schieltz D, Yates III JR, Grant
PA . Chd1 chromodomain links histone H3 methylation with SAGA- and SLIK-dependent acetylation. _Nature_ 2005; 433:434–438. Article CAS PubMed Google Scholar * Taverna SD, Ilin S, Rogers
RS, _et al_. Yng1 PHD finger binding to H3 trimethylated at K4 promotes NuA3 HAT activity at K14 of H3 and transcription at a subset of targeted ORFs. _Mol Cell_ 2006; 24:785–796. Article
CAS PubMed PubMed Central Google Scholar * Kim J, Guermah M, McGinty RK, _et al_. RAD6-mediated transcription-coupled H2B ubiquitylation directly stimulates H3K4 methylation in human
cells. _Cell_ 2009; 137:459–471. Article CAS PubMed PubMed Central Google Scholar * Lee JS, Shukla A, Schneider J, _et al_. Histone crosstalk between H2B monoubiquitination and H3
methylation mediated by COMPASS. _Cell_ 2007; 131:1084–1096. Article CAS PubMed Google Scholar * Zippo A, Serafini R, Rocchigiani M, Pennacchini S, Krepelova A, Oliviero S . Histone
crosstalk between H3S10ph and H4K16ac generates a histone code that mediates transcription elongation. _Cell_ 2009; 138:1122–1136. Article CAS PubMed Google Scholar * Guccione E, Bassi
C, Casadio F, _et al_. Methylation of histone H3R2 by PRMT6 and H3K4 by an MLL complex are mutually exclusive. _Nature_ 2007; 449:933–937. Article CAS PubMed Google Scholar * Wu Y, Li Q,
Chen XZ . Detecting protein-protein interactions by Far western blotting. _Nat Protoc_ 2007; 2:3278–3284. Article CAS PubMed Google Scholar * Brenner S . The genetics of Caenorhabditis
elegans. _Genetics_ 1974; 77:71–94. CAS PubMed PubMed Central Google Scholar * Yang Y, Hu L, Wang P, _et al_. Structural insights into a dual-specificity histone demethylase ceKDM7A from
_Caenorhabditis elegans_. _Cell Res_ 2010; 20:886–898. Article CAS PubMed Google Scholar Download references ACKNOWLEDGEMENTS We thank Yang Shi (Harvard Medical School, USA) and Ruiming
Xu (Institute of Biophysics, CAS, China) for their critical reading of the manuscript, the National BioResource Project for _C. elegans_ (Japan) for the F29B9.2 (tm3713) mutant strain, the
cell biology and molecular biology core facilities for MS and biacore experiments, and other members in Chen lab for technical help. This work was supported by grants from the National Basic
Research Program of China (2010CB529700, 2009CB918600, and 2007CB947900), the National Natural Science Foundation of China (30870493, 90919026), Chinese Academy of Sciences (KSCX2-YW-R-04),
Shanghai Pujiang Program (08PJ14010, 0757S11361), Shanghai Leading Academic Discipline Project (B111), and the Council of Shanghai Municipal Government for Science and Technology. AUTHOR
INFORMATION AUTHORS AND AFFILIATIONS * State Key Laboratory of Molecular Biology, Shanghai Key Laboratory of Molecular Andrology, Institute of Biochemistry and Cell Biology, Shanghai
Institutes for Biological Sciences, Chinese Academy of Sciences, 320 Yueyang Road, Shanghai, 200031, China Hanqing Lin, Yiqin Wang, Yanru Wang, Feng Tian, Pu Pu, Yi Yu, Hailei Mao &
Charlie Degui Chen * Affiliated Hospital of Nantong University, 226001, Nantong, China Yanru Wang & Hailei Mao * School of Life Sciences, Fudan University, 220 Han-Dan Road, Shanghai,
200433, China Ying Yang, Ping Wang, Lulu Hu, Yan Lin, Yi Liu & Yanhui Xu * Institutes of Biomedical Sciences, Fudan University, 130 Dong-An Road, Shanghai, 200032, China Ying Yang, Ping
Wang, Lulu Hu, Yan Lin, Yi Liu & Yanhui Xu Authors * Hanqing Lin View author publications You can also search for this author inPubMed Google Scholar * Yiqin Wang View author
publications You can also search for this author inPubMed Google Scholar * Yanru Wang View author publications You can also search for this author inPubMed Google Scholar * Feng Tian View
author publications You can also search for this author inPubMed Google Scholar * Pu Pu View author publications You can also search for this author inPubMed Google Scholar * Yi Yu View
author publications You can also search for this author inPubMed Google Scholar * Hailei Mao View author publications You can also search for this author inPubMed Google Scholar * Ying Yang
View author publications You can also search for this author inPubMed Google Scholar * Ping Wang View author publications You can also search for this author inPubMed Google Scholar * Lulu
Hu View author publications You can also search for this author inPubMed Google Scholar * Yan Lin View author publications You can also search for this author inPubMed Google Scholar * Yi
Liu View author publications You can also search for this author inPubMed Google Scholar * Yanhui Xu View author publications You can also search for this author inPubMed Google Scholar *
Charlie Degui Chen View author publications You can also search for this author inPubMed Google Scholar CORRESPONDING AUTHORS Correspondence to Yanhui Xu or Charlie Degui Chen. ADDITIONAL
INFORMATION ( SUPPLEMENTARY INFORMATION is linked to the online version of the paper on the _Cell Research_ website.) SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION, FIGURE S1 Mutation
of H495 abolished the demethylase activity. (PDF 31 kb) SUPPLEMENTARY INFORMATION, FIGURE S2 Venn diagram of ceKDM7A and H3K4me3 bound genes identified by ChIP-Seq with the cutoff of one
sequence tag. (PDF 12 kb) SUPPLEMENTARY INFORMATION, TABLE S1 Q-PCR analysis of gene expression comparing ceKDM7A knockout Tm3713 to wildtype N2. (PDF 21 kb) RIGHTS AND PERMISSIONS Reprints
and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Lin, H., Wang, Y., Wang, Y. _et al._ Coordinated regulation of active and repressive histone methylations by a dual-specificity histone
demethylase ceKDM7A from _Caenorhabditis elegans_. _Cell Res_ 20, 899–907 (2010). https://doi.org/10.1038/cr.2010.84 Download citation * Received: 23 March 2010 * Revised: 04 May 2010 *
Accepted: 05 May 2010 * Published: 22 June 2010 * Issue Date: August 2010 * DOI: https://doi.org/10.1038/cr.2010.84 SHARE THIS ARTICLE Anyone you share the following link with will be able
to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing
initiative KEYWORDS * KDM7A * demethylase * H3K9me2 * H3K27me2 * PHD * JmjC * histone * methylation