The aglycone of ginsenoside rg3 enables glucagon-like peptide-1 secretion in enteroendocrine cells and alleviates hyperglycemia in type 2 diabetic mice
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT Ginsenosides can be classified on the basis of the skeleton of their aglycones. Here, we hypothesized that the sugar moieties attached to the dammarane backbone enable binding of
the ginsenosides to the sweet taste receptor, eliciting glucagon-like peptide-1 (GLP-1) secretion in the enteroendocrine L cells. Using the human enteroendocrine NCI-H716 cells, we
demonstrated that 15 ginsenosides stimulate GLP-1 secretion according to the position of their sugar moieties. Through a pharmacological approach and RNA interference technique to inhibit
the cellular signal cascade and using the Gαgust−/− mice, we elucidated that GLP-1 secreting effect of Rg3 mediated by the sweet taste receptor mediated the signaling pathway. Rg3, a
ginsenoside metabolite that transformed the structure through a steaming process, showed the strongest GLP-1 secreting effects in NCI-H716 cells and also showed an anti-hyperglycemic effect
on a type 2 diabetic mouse model through increased plasma GLP-1 and plasma insulin levels during an oral glucose tolerance test. Our study reveals a novel mechanism where the sugar moieties
of ginsenosides Rg3 stimulates GLP-1 secretion in enteroendocrine L cells through a sweet taste receptor-mediated signal transduction pathway and thus has an anti-hyperglycemic effect on the
type 2 diabetic mouse model. SIMILAR CONTENT BEING VIEWED BY OTHERS CYANIDIN-3-O-GLUCOSIDE ENHANCES GLP-1 SECRETION VIA PPARΒ/Δ-Β-CATENIN-TCF-4 PATHWAY IN TYPE 2 DIABETES MELLITUS Article
Open access 20 May 2025 THE ANTIDIABETIC EFFECT OF SAFFLOWER YELLOW BY REGULATING THE GOAT/GHRELIN/GHS-R1A/CAMP/TRPM2 PATHWAY Article Open access 11 February 2025 MOGROL STIMULATES
G-PROTEIN-COUPLED BILE ACID RECEPTOR 1 (GPBAR1/TGR5) AND INSULIN SECRETION FROM PANCREATIC Β-CELLS AND ALLEVIATES HYPERGLYCEMIA IN MICE Article Open access 08 February 2024 INTRODUCTION
Ginsenosides are triterpenoid saponins found abundantly in Panax species (ginseng). The dammarane type family consists mostly of ginsenosides and is largely composed of a four-carbon ring
structure backbone with various sugar moieties attached to the C-3 and C-20 positions and can be divided into three main groups: protopanaxadiols (PPDs), protopanaxatriols (PPTs) and
ocotillol1,2. PPTs, such as Re, Rf, Rg1 and Rh1, are composed of sugar moieties attached to the α-OH at C-6 and/or β-OH at C-20 of the dammarane skeleton, while PPDs, such as Rb1, Rb2, Rc,
Rd, Rg3 and Rh2, are composed of sugar moieties attached to the β-OH at C-3 or C-203. Most ginsenosides have been isolated from roots, leaves, stems, fruits, and/or flower parts of _Panax
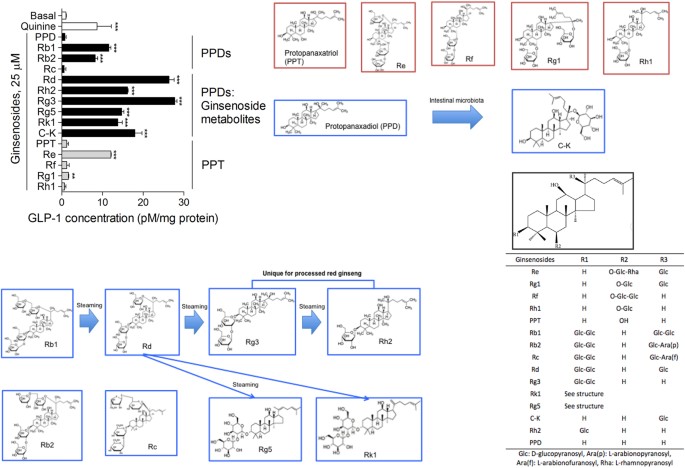
ginseng_, but several types can be converted into other types of ginsenosides by a steaming process or intestinal microbiota metabolism2. For example, the ginsenoside Rg3, which is absent in
raw ginseng, is a bioactive compound of red ginseng, a heat processed _Panax ginseng_ and PPDs, such as Rb1, Rb2, Rc and Rd, can be converted into compound K (C-K) by the intestinal
microbiota metabolism4,5. The antidiabetic and antiobesity effects of several ginsenosides including Rb1, Re, Rb2, Rg3 and C-K have been investigated5,6,7,8,9. We focused on the structure of
ginsenosides, which contain various sugar moieties attached to the dammarane skeleton and hypothesized that the sugar moieties stimulate GLP-1 secretion via activation of a sweet taste
receptor signaling pathway in enteroendocrine L cells. GLP-1 is a potent anti-hyperglycemic agent, which induces glucose-dependent insulin secretion from pancreatic β cells, while suppresses
glucagon secretion. For intact GLP-1 in plasma removed rapidly by renal clearance and enzyme dipeptidyl peptidase-4 (DPP-4), incretin-based treatment using DPP-4 inhibitors (gliptins) or
GLP-1 receptor (GLP-1R) agonists is a frequently prescribed therapy for type 2 diabetes mellitus10,11. The NCI-H716 cell line is derived from ascetic fluid of a 33 year old Caucasian male
patient with a poorly differentiated cecal adenocalcinoma12. Culturing the cells with a specific extracellular matrix causes endocrine differentiation, leading the cells to express several
neuroendocrine markers including chromogranin A and on this basis they are a qualified enteroendocrine cellular model for studying the regulation of GLP-1 secretion12. We have reported that
enteroendocrine L cells express taste receptors and their downstream signal elements, including a specific G protein, Gα-gustducin (Gαgust) and G protein-coupled sweet and bitter taste
receptors, similar to their expression in the tongue13,14. T1R3 taste receptor, which is expressed by about 10-20% of taste cells, consists of a heterodimer T1R2 that recognizes a broad
spectrum of sweet taste stimuli, including natural and synthetic sugar15,16. Previous reports suggest that the intracellular signal transduction pathway activated by sugar binding to taste
receptors is mediated by the activation of Gαgust and a consequent signaling cascade including phospholipase Cβ2 (PLCβ2) and inositol 1,4,5-triphosphate (IP3)16,17. This Gβγ-subunit
mediating the signaling cascade elicits the release of Ca2+ from intracellular stores and subsequent Ca2+-dependent activation of a transient receptor potential channel M5 (TRPM5), leading
to the membrane depolarization and further action potential generation in turn18,19. In this study, we demonstrated the GLP-1 secreting effect of ginsenosides using the enteroendocrine
NCI-H716 cell line. Rg3, a PPD group ginsenoside that is abundant in steamed ginseng, showed the strongest GLP-1 secreting effect. Using the cell line and Gαgust−/− mice, we investigated the
cellular mechanism underlying the GLP-1 secreting effect of Rg3 and using _db/db_ mice, we evaluated the possibility of exploiting the effect of Rg3 as a therapeutic agent for type 2
diabetes mellitus. RESULTS To confirm our hypothesis that the ginsenosides stimulate GLP-1 secretion in enteroendocrine L cells, we treated endocrine differentiated NCI-H716 cells with 15
types of ginsenosides. We treated the PPD family, PPD, Rb1, Rb2, Rc, Rd, Rh2, Rg3, Rg5, Rk1 and C-K and the PPT family, PPT, Re, Rf, Rg1 and Rh12. We observed GLP-1 secretion in NCI-H716
cells treated with ginsenosides: Rb1, Rb2, Rd, Rg3, Rg5, Rk1, C-K, Re and Rg1 (Fig. 1). Interestingly, Rd, Rg3, Rh2, Rg5 and Rk1, which are nearly absent in _Panax ginseng_, but abundantly
found in red ginseng, showed a strong GLP-1 secreting effect in the NCI-H716 cells. The ginsenoside metabolites appear to result from deglycosylation during the steaming process1. The most
abundantly found ginsenosides in _Panax ginseng_ are Rb1, Rb2, Rc, Re and Rg1. We observed a GLP-1 secreting effect of Rb1, Rb2, Re and Rg1 on NCI-H716 cells, but Rc, Rf and Rh1 had no GLP-1
secreting effect on the cells (Fig. 1). C-K, a particular ginsenoside that is only produced by intestinal microbiota metabolism, strongly stimulated GLP-1 secretion in the cells (Fig. 1).
We also treated the cells with PPD and PPT but no GLP-1 secreting effect was observed (Fig. 1). The GLP-1 secreting effects of ginsenosides were compared to the effect of the bitter tastant
quinine20. Ginsenoside Rg3 showed the strongest GLP-1 secreting effect in the NCI-H716 cells (Fig. 1). In this study, we performed further _in vitro_ and _in vivo_ studies using Rg3. We
observed a dose-dependent GLP-1 secreting effect of Rg3 treatment in NCI-H716 cells (Fig. 2a). Rg3 did not affect the cell’s viability below a concentration of 25 μM (Fig. 2b). To examine
whether the GLP-1 secreting effect of Rg3 is mediated by sweet taste receptor activation, we transfected siRNAs targeting the _T1R2_ and/or _T1R3_ to the NCI-H716 cells and measured the
GLP-1 levels stimulated by Rg3 treatment (Fig. 2c). We also measured glucose stimulated GLP-1 secretion to confirm our RNA interference sets (Fig. 2d). Rg3 (10 μM) showed 2-fold GLP-1
secreting effect compare to the glucose (10%, w/v). One of the _T1R2_ or _T1R3_ siRNA transfection partly decreased the GLP-1 secreting effects of Rg3 and glucose, respectively (Fig. 2c,d).
Glucose-stimulated GLP-1 secretion was completely blocked by the _T1R2_ and _T1R3_ double siRNA transfection while Rg3-stimulated GLP-1 secretion slightly remained. The siRNAs accurately
knocked-down their targeting mRNA expressions (Fig. 2e). A human sweet taste receptor antagonist lactisole, which have reported to block T1R3, was pre-treated NCI-H716 cells and
significantly inhibited Rg3 stimulated GLP-1 secretion (Fig. 2f)21. We tried to identify the cellular downstream pathway of Rg3 stimulated GLP-1 secretion in the NCI-H716 cells. We blocked
several GPCRs and Gαgust by transfecting siRNAs targeting the signaling molecules. Using siRNAs targeting the Gαgust gene, _GNAT3_, a cannabinoid receptor gene, _GPR119_ and a bile acid
receptor 1 gene, _GPBAR1_ (aliases _TGR5_), we found the GLP-1 secreting effect of Rg3 is mediated by Gαgust (Fig. 3a). We confirmed our RNA interference sets with C-K (Fig. 3b), which was
reported to stimulate bile acid receptor-mediated GLP-1 secretion5 and also with denatonium benzoate (DB) (Fig. 3c), which was reported to stimulate Gαgust-mediated GLP-1 secretion14. The
siRNAs accurately knocked-down their targeting mRNA expressions (Fig. 3d). We then, traced the intracellular signaling pathway of Rg3 stimulated GLP-1 secretion in NCI-H716 cells. Using
pathway inhibitors, Gβγ inhibitor gallein (Fig. 4a), PLC inhibitor U73122 (Fig. 4b), IP3 receptor antagonist 2-aminoethoxydiphenyl borate (2APB) (Fig. 4c) and PKC inhibitor
bisindolylmaleimide I (BIM) (Fig. 4d), we demonstrated that the GLP-1 secreting effect of Rg3 is mediated by Gβγ-PLC-IP3 signaling elements, similar to the sweet taste receptor in the
tongue, in the enteroendocrine NCI-H716 cells. We also measured [Ca2+]i levels in the NCI-H716 cells after Rg3 treatment. Enteroendocrine NCI-H716 cells were treated with Rg3 without
extracellular calcium. Rg3 treatment elicited [Ca2+]i release from the intracellular calcium store, the endoplasmic reticulum (ER) and the effect was absent in the cells that were
pre-treated with the sweet taste inhibitor lactisole (Fig. 4e). We have measured GLP-1 secreting effect of acesulfame K, a well-known artificial sweetener, on the NCI-H716 cells. However,
Acesulfame K treatment did not affect the GLP-1 secreting effect of the NCI-H716 cells (Fig. 4f). We also traced the physiological role of the Gα subunit, which has not been extensively
considered in sweet taste receptor research, during Rg3 treatment. A common Gαgust-mediated signaling pathway response to the activation of G protein-coupled taste receptors, such as T1Rs or
T2Rs, involves activation of phosphodiesterase (PDE) and continuous intracellular cAMP reduction14,16,22. Surprisingly, we observed that the GLP-1 secreting effect of Rg3 is mediated by AC
activation rather than PDE. Enteroendocrine NCI-H716 cells that were pre-treated with the AC inhibitor SQ22536 (Fig. 5a) or protein kinase A (PKA) inhibitor H89 (Fig. 5b) inhibited the GLP-1
secreting effect of Rg3, whereas the pan-PDE inhibitor 3-isobutyl-1-methylxanthine (IBMX) (Fig. 5c) did not affect the GLP-1 secreting effect. We measured the Rg3 stimulated intracellular
cAMP production in the NCI-H716 cells by 15 min intervals. A large amount of intracellular cAMP was measured at 15 min after Rg3 treatment and was lowered to the basal level after 45 min
(Fig. 5d). The stimulatory effect of Rg3 treatment on the intracellular cAMP production was abolished in the lactisole pre-treated cells (Fig. 5d). We then examined the phosphorylation of
cAMP-dependently activated signal transduction elements. PKA, a cAMP-dependent protein kinase, CREB, a cAMP-responsive transcription factor and extracellular signal regulated kinase (ERK)
1/2, a regulator of CREB function, were phosphorylated after Rg3 treatment (Fig. 5e). We provide a figure that suggests the intracellular signal transduction event mediated by sweet taste
receptor activation in NCI-H716 cells upon the Rg3 treatment (Fig. 6). Subsequently, we performed oral glucose tolerance test (OGTT) using _db/db_ mice and observed the effect of oral
administration of Rg3 on hyperglycemia. The fasting glucose levels of _db/db_ mice ranged from 200–300 mg/dl. Comparing to the saline-treated _db/db_ mouse group, the Rg3-treated group
showed a delayed increase in blood glucose level at 20 min after the treatment and showed lowered blood glucose levels at 90 and 120 min after the treatment (Fig. 7a). We also performed OGTT
with the same _db/db_ mouse groups and collected the blood to measure the effect of Rg3 administration on the plasma GLP-1 and plasma insulin levels. Fasting plasma GLP-1 levels of _db/db_
mice ranged from 70–190 pg/ml and the plasma GLP-1 level in the Rg3-treated mouse group was increased about 2-fold after 10 min during the OGTT (Fig. 7b). Fasting plasma insulin levels
ranged from 4793–12982 pg/ml and the Rg3 treatment also increased the plasma insulin level about 1.3-fold after 10 min during the OGTT (Fig. 7c). The area under the curve shows the variation
in the blood glucose levels of each _db/db_ mouse group during OGTT (Fig. 7d). We performed the same experiments using Gαgust−/− mice to elucidate the involvement of Gαgust in the
anti-hyperglycemic effect of Rg3 administration. We observed lowered blood glucose levels in both C57- and Gαgust−/− mouse groups after 20 min of Rg3 administration during OGTT (Fig. 8a,b).
However, the increased plasma GLP-1 level observed in the Rg3-treated C57 mouse group (Fig. 8c) was abolished in the Gαgust−/− mouse group during the OGTT (Fig. 8d). Moreover, the
stimulatory effect of Rg3 administration observed in the Rg3-treated C57 mouse group (Fig. 8e) was also abolished in the Gαgust−/− mouse group at the same time (Fig. 8f). DISCUSSION _Panax
ginseng_ is considered one of the most valuable medicinal plants in Asia and is largely consumed throughout the world. Ginsenosides, the saponins found in all parts of the ginseng plant,
have been investigated for their various pharmacological effects on hyperglycemia, weight gain, neuroprotection, tumor cell growth and hypertension. Regarding the diverse pharmacological
aspects of ginsenosides, Attle _et al._ explained that the ginsenosides share structural similarities with steroid hormones, especially progesterone and pregnanolone and thereby have
numerous physiological activities23. Structural diversity including type-, number- and site of attachment of sugar moieties also contributes to the diverse pharmacological effects of
ginsenosides23. This structural diversity can be amplified through a steaming process and intestinal microbiota metabolism. We focused on the chemical structure of dammarane family
ginsenosides having various sugar moieties with their carbon-ring backbone and hypothesized that these sugar moieties act like ligands for the sweet taste receptor. We demonstrated the GLP-1
secreting effect of 15-dammarane family ginsenosides in human enteroendocrine NCI-H716 cells. PPDs that have sugar moieties attached at C-3 and C-20 showed a GLP-1 secreting effect on the
NCI-H716 cells. On the other hand, PPTs that have sugar moieties attached at C-6 did not show the GLP-1 secreting effect on the NCI-H716 cells while the ginsenoside Re and Rg1, which has a
glucose residue at C-20, showed a moderate GLP-1 secreting effect. From the obtained results, we assume that the sugar moieties attached at the C-3 and/or C-20 contribute to the binding
affinity of the dammarane family ginsenosides to the sweet taste receptor. Interestingly, ginsenoside metabolites, the chemical structure of which was transformed through a steaming process
or intestinal microbiota metabolism, showed the strongest GLP-1 secreting effect. Ginsenoside Rg3 showed the strongest GLP-1 secreting effect and we traced the intracellular mechanism using
enteroendocrine NCI-H716 cells. Structurally, Rg3 is a PPD with two D-glycopyranosyl moieties. Thus we have assumed that the sugar moieties of ginsenoside Rg3 provide a binding motif to the
sweet taste receptor expressed on the enteroendocrine L cells. GLP-1 secreting effect of Rg3 was significantly decreased in each T1R2 or T1R3 siRNA transfected NCI-H716 cells, respectively.
But the effect was not removed completely even both the siRNAs transfection. A considerable GLP-1 secretion in the both siRNAs transfected cells response to Rg3 stimuli suggesting the
existence of multiple receptors for Rg3. A recent study reported that a ginsenoside metabolite C-K stimulates GLP-1 secretion in NCI-H716 cells via binding of a bile acid receptor5. We
assumed that the dammarane backbone of Rg3 may contribute a binding motif to the other receptors, such as a bile acid receptor or a cannabinoid receptor and transfected the corresponding
siRNAs to abolish the mRNA expression. The GLP-1 secreting effect of Rg3 was not mediated by the cannabinoid- or bile acid receptors. Nevertheless, the GLP-1 secreting effect of Rg3 appears
to be dependent on the Gαgust. Therefore, we assume participation of one or more bitter taste receptor activation in the GLP-1 secreting effect of Rg3 alongside sweet taste receptor. We
traced a common sweet taste modulatory cellular pathway, which is mediated by the Gβ3γ13-PLCβ2-IP3 signal cascade. Through pharmacological approaches using corresponding inhibitors or
antagonist, we determined that the GLP-1 secreting effect of Rg3 is mediated by the signal cascade, but also found that a considerable GLP-1 response remained. Gαgust is expected to cause
activation of PDE and thus decrease intracellular cAMP levels24. Indeed, we observed that the bitter tastant DB decreased intracellular cAMP levels during its GLP-1 secreting event
responding to Gαgust activation14. However, similar to studies showing that sugars increase intracellular cAMP levels, we also have found that Rg3 increases intracellular cAMP levels. We
further showed the involvement of enzyme AC, which produces cAMP in response to the stimuli, in the GLP-1 secreting effect of Rg3 instead of PDE. Moreover, phosphorylations of PKA and CREB,
which are activated in response to the increased intracellular cAMP level, are involved in the GLP-1 secreting effect of Rg3. PKA is also involved in the GLP-1 secreting effect of Rg3.
ERK1/2, a mitogen-activated protein kinase (MAPK), is known to activate various transcription factors including CREB, in response to diverse extracellular stimuli such as forskolin25. One of
the interesting results in our study is that acesulfame K, an artificial sugar, did not affect the GLP-1 secreting effect of NCI-H716 cells. Since the expression of sweet taste receptors
have been found in the enteroendocrine L cells along with its signal transduction elements artificial sugars had been convinced that they are able to activate the sweet taste receptors in
the L cells as they do in the lingual tissues13. In contrast to the _in vitro_ studies that show GLP-1 secreting effect of artificial sugars in the human and mouse enteroendocrine cells, _in
vivo_ studies using healthy human subjects failed to show the effects of artificial sugars on the GLP-1 secretion26,27,28. However, a human study demonstrated that the sweet taste receptor
inhibitor drastically blocked the GLP-1 and PYY secreting effect of glucose29. Perhaps, the GLP-1 and PYY secreting effect via activation of sweet taste receptor in the enteroendocrine L
cells depends on the structural analogy to glucose than the sweetness itself. Our _in vivo_ study elucidated the effects of ginsenoside Rg3 administration on hyperglycemia in type 2 diabetic
mice. The therapeutic effects of Rg3 against metabolic disorder, such as obesity and hyperglycemia have been demonstrated. Park _et al._ reported enhanced glucose-stimulated insulin
secretion via AMP-activated protein kinase (AMPK) activation upon Rg3 treatment in a hamster pancreatic β cell line7. The effects of Rg3 on AMPK activation suppressed adipocyte
differentiation in mouse 3T3-L1 adipocyte and improved glucose uptake in the rat L6 myocyte30,31. Therefore, Rg3 has direct glucose lowering effect on hyperglycemia through enhanced glucose
uptake activity in the myoblast and also has an indirect effect through stimulation of insulin secretion in the pancreatic β cell. In this study, Rg3 stimulated GLP-1 secretion by activating
sweet taste receptor-and Gαgust-mediated signal transduction cascade in enteroendocrine cells and increased plasma GLP-1 and plasma insulin levels in _db/db_ mice after glucose gavage.
Using Gαgust−/− mice, we showed the stimulatory effect of Rg3 on plasma GLP-1 and that plasma insulin is Gαgust-dependent. The slightly lowered blood glucose level after Rg3 administration
observed in the Gαgust−/− mice appears to demonstrate a direct effect of Rg3 on the blood glucose regulation7. Our study elucidates a novel mechanism underlying the anti-diabetic effects of
ginsenoside Rg3. The sugar moieties of Rg3 and other dammarane family ginsenosides enables their binding to the sweet taste receptor to stimulate GLP-1 secretion in intestinal L cells. The
GLP-1 secreting effect lowers the blood glucose levels through insulinotropic action. Considering the diverse benefits of GLP-1 on hyperglycemia, food intake and even β cell function, Rg3
has possibility to be developed as a novel therapeutic agent for type 2 diabetes and obesity. METHODS CHEMICALS All ginsenosides were kindly provided by Dr. Kwang-Seok Ahn (Department of
Pathology, College of Korean Medicine, Kyung Hee University, Seoul, South Korea). Rg3, DB, metformin hydrochloride, acesulfame K, D-glucose, forskolin and the GPCR pathway inhibitors;
U73122, BIM, H-89, IBMX were purchased from Sigma-Aldrich (St. Louis, MO, USA). Gallein, 2APB and SQ22536 were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Lactisole was
purchased from Endeavour Speciality Chemicals (Daventry, UK). CELL CULTURE Human NCI-H716 cells were obtained from the Korean Cell Line Bank (KCLB®, Seoul, South Korea). Cells were
maintained in RPMI 1640 (Lonza, Walkersville, MD, USA) supplemented with 10% fetal bovine serum (FBS; Lonza). For endocrine differentiation, the cells were plated in matrigel (BD Bioscience,
Bedford, MA, USA)-precoated 24-well plates at 5 × 105 cells per well. The cells were incubated for 48 h in a humidified CO2 incubator as described previously13,14. GLP-1 ELISA The endocrine
differentiated cell media was replaced with PBS containing 1 mM calcium chloride and different drug concentrations. The GPCR pathway inhibitors were pre-incubated for 30 min before the drug
treatment. After incubation for 1 h in a CO2 incubator, GLP-1 concentration was measured as previously described14. An active GLP-1 ELISA (EMD Millipore, Billerica, MA, USA) was performed
as described in the manufacturer’s instructions. The active GLP-1 concentrations in each sample were measured using a Fluoroskan Ascent FL machine (Thermo Fishwer Scientific, Vantaa,
Finland). The lowest level of active GLP-1 that can be detected by the GLP-1 assay is 2 pM. CELL VIABILITY ASSAY A cell viability assay was performed using
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. SIRNA TRANSFECTION siRNA duplexes for _T1R2_,
_T1R3_, _GNAT3_ (Gαgust gene), _GPR119_ and _GPBAR1_ were synthesized by Bioneer (Bioneer Co., Daejeon, South Korea). The information for each siRNA was provided online (Supplementary Table
S1). A scrambled negative control siRNA was purchased from Bioneer. Endocrine differentiated NCI-H716 cells were transfected with the siRNA duplexes using Lipofectamine RNAiMAX reagent
(Invitrogen). REAL-TIME QUANTITATIVE PCR The expression of _T1R2_, _T1R3_, _GNAT3_, _GPR119_ and _GPBAR1_ after siRNA transfection was determined using a StepOne real-time PCR instrument
(Applied Biosystems, Foster City, CA, USA). Total RNA isolation and subsequent cDNA hybridization were performed as previously described32. The expression levels of _T1R2_, _T1R3_, _GNAT3_,
_GPR119_ and _GPBAR1_ in each type of siRNA-transfected cell were compared with the corresponding levels in the negative control siRNA-transfected cells and the the 2−ΔΔCt values were
determined33. _GAPDH_ was used as an endogenous control. The sense and antisense sequence of each primer were provided online (Supplementary 02). CALCIUM IMAGING NCI-H716 cells were seeded
on a clear-bottom 96-well black plate (Corning, Tewksbury, MA, USA). After differentiation, the medium was replaced with PBS and the mixture was incubated for 30 min with fura-2 AM dye as
described previously14,34. After 30 min, the medium was replaced with saline with or without 2.5 mM lactisole and incubated for a further 30 min. [Ca2+]i were observed with a Nikon Eclipse
TS 100 fluorescence imaging system (Nikon, Melville, NY, USA) and quantified and visualized with InCyt Im2 software (University of Cincinnati, Cincinnati, OH, USA). The number of cells
observed was 10–20 per well. CAMP ELISA Endocrine differentiated NCI-H716 cells were incubated with Rg3 or forskolin. The drug-treated cells were collected at 15 min intervals. Lactisole was
pre-treated prior to Rg3 treatment. The collected cells were lysed using 0.1 M HCl and the intracellular cAMP was assayed using ELISA (Enzo Life Sciences, Farmingdale, NY, USA) according to
the manufacturer’s instructions. The results were normalized to the protein concentration. IMMUNOBLOTING Cells were lysed with cell lysis buffer (1% NP-40, 150 mM NaCl, 20 mM Tris-HCl, 1 mM
EDTA, 1 mM EGTA and a proteinase inhibitor cocktail). Total protein (5 μg) was loaded into each well and immunoblotting was performed as described previously13,14,35. Individual proteins
were detected with primary antibodies against the phosphorylated form of PKA-C (1:1000 dilution), total PKA-C (1:1,000 dilutions), the phosphorylated form of cAMP-response element-binding
protein (CREB) (1:3,000 dilution), total CREB (1:3,000 dilution), the phosphorylated form of ERK 1/2 (1: 3,000 dilution) and total ERK 1/2 (1:3,000 dilution). All primary antibodies were
purchased from Cell Signaling (Danvers, MA, USA). After incubation with horseradish peroxidase conjugated secondary antibodies (Santa Cruz Biotechnologies), signals were visualized by
enhanced chemiluminescence (Daeil Lab Service Co., Ltd. Seoul, South Korea). ANIMALS All experiments were performed in accordance with the guidelines and the regulations of the Institutional
Ethical Committee of Kyung Hee University Institutional Animal Care and Use. All animal study protocols were approved by the Institutional Animal Care and Use Committee of Kyung Hee
University (KHUASP(SE)-12-032). Eight-week old male _db/db_ mice and C57BL/6 mice were purchased from Daehan Biolink (DBL, Eumseoung-gun, Chungcheongbuk-do, South Korea). The original mating
pairs of Gαgust−/− mice were kindly provided by Dr. Robert F. Margolskee (Monell Chemical Senses Center, Philadelphia, PA, USA). Gαgust−/− mice, a homozygous null for the _Gnat3_ allele,
were created using homologous recombination in 129/Sv background embryonic stem (W9.5) cells and genotyped as previously described36. All animals were acclimated for one week before the
experiment. The animals were housed in a room with a light-dark cycle of 12 h at a temperature ranging from 21–23 °C and moderate humidity (55–60%). Food and water were provided _ad
libitum_. OGTT Mice were fasted for 18 h before the OGTT. Each mouse group was orally administered saline or Rg3 (0.5 mg/Kg) just before glucose gavage (5 g/Kg). The blood glucose was
measured from the tail vein using an Accu-Check Performa system (Roche Diagnostics, Mannheim, Germany) at 6 time points: 0 (before glucose gavage), 10 (after glucose gavage), 20, 40, 90 and
120 min. PLASMA GLP-1 AND PLASMA INSULIN ASSAY To prevent hypotensive shock, all mice were allowed to rest for one week. Mice were fasted for 18 h before the experiments. Each mouse group
was orally administered saline or Rg3 (0.5 mg/Kg) just before the glucose gavage (2 g/Kg). The collected blood from the tail veins was immediately transferred into EDTA-coated
microcentrifuge tubes containing a dipiptidyl peptidase IV inhibitor (EMD Millipore) and a protease inhibitor cocktail (Roche Diagnostics). The collected blood samples were centrifuged at
1,000 × _g_ for 20 min at 4 °C and the plasma was carefully separated into fresh tubes. A multiplex assay (Mouse Diabetes panel: total GLP-1 and insulin; Bio-Rad) was performed as described
in the manufacturer’s instructions. The total GLP-1 and insulin concentrations in each sample were measured using a Bio-Plex MAGPIX Multiplex reader (Bio-Rad). The results were analyzed with
Bio-Plex Manager software (Bio-Rad). STATISTICAL ANALYSIS GraphPad Prism 5 software (GraphPad Software, San Diego, CA, USA) was used for the statistical analysis of the experimental
results. The results from the GLP-1 and cAMP ELISA represent at least three separate experiments performed in quadruplicate. The statistical significance of each ELISA and area under the
curve (AUC) graph was measured using Mann-Whitney U test (one-tailed) or a one-way ANOVA with Bonferroni’s post hoc test. For OGTT and the mouse plasma hormone studies, Mann-Whitney U test
was done for the inter group comparison at each time point. Five to eight mice per group were used for the _in vivo_ studies. The _in vivo_ studies were performed at least twice. ADDITIONAL
INFORMATION HOW TO CITE THIS ARTICLE: Kim, K.-S. _et al._ The aglycone of ginsenoside Rg3 enables glucagon-like peptide-1 secretion in enteroendocrine cells and alleviates hyperglycemia in
type 2 diabetic mice. _Sci. Rep._ 5, 18325; doi: 10.1038/srep18325 (2015). REFERENCES * Leung, K. W. & Wong, A. S. Pharmacology of ginsenosides: a literature review. Chin Med 5, 20
(2010). Article PubMed PubMed Central Google Scholar * Christensen, L. P. Ginsenosides chemistry, biosynthesis, analysis and potential health effects. Adv Food Nutr Res 55, 1–99 (2009).
CAS PubMed Google Scholar * Kim, Y. J., Zhang, D. & Yang, D. C. Biosynthesis and biotechnological production of ginsenosides. Biotechnol Adv 33, 717–735 (2015). Article CAS PubMed
Google Scholar * Jo, S. K., Kim, I. S., Yoon, K. S., Yoon, H. H. & Yoo, H. H. Preparation of ginsenosides Rg3, Rk1 and Rg5-selectively enriched ginsengs by a simple steaming process.
European Food Research and Technology 240, 251–256 (2015). Article CAS Google Scholar * Kim, K., Park, M., Lee, Y. M., Rhyu, M. R. & Kim, H. Y. Ginsenoside metabolite compound K
stimulates glucagon-like peptide-1 secretion in NCI-H716 cells via bile acid receptor activation. Arch Pharm Res 37, 1193–1200 (2014). Article CAS PubMed Google Scholar * Lee, K. T. et
al. The antidiabetic effect of ginsenoside Rb2 via activation of AMPK. Arch Pharm Res 34, 1201–1208 (2011). Article CAS PubMed Google Scholar * Park, M. W., Ha, J. & Chung, S. H.
20(S)-ginsenoside Rg3 enhances glucose-stimulated insulin secretion and activates AMPK. Biol Pharm Bull 31, 748–751 (2008). Article CAS PubMed Google Scholar * Xie, J. T. et al.
Anti-diabetic effect of ginsenoside Re in ob/ob mice. Biochim Biophys Acta 1740, 319–325 (2005). Article CAS PubMed Google Scholar * Xiong, Y. et al. Antiobesity and antihyperglycemic
effects of ginsenoside Rb1 in rats. Diabetes 59, 2505–2512 (2010). Article CAS PubMed PubMed Central Google Scholar * Drucker, D. J. et al. Incretin-based therapies for the treatment of
type 2 diabetes: evaluation of the risks and benefits. Diabetes Care 33, 428–433 (2010). Article CAS PubMed PubMed Central Google Scholar * Holst, J. J. The physiology of glucagon-like
peptide 1. Physiol Rev 87, 1409–1439 (2007). Article CAS PubMed Google Scholar * Reimer, R. A. et al. A human cellular model for studying the regulation of glucagon-like peptide-1
secretion. Endocrinology 142, 4522–4528 (2001). Article CAS PubMed Google Scholar * Jang, H. J. et al. Gut-expressed gustducin and taste receptors regulate secretion of glucagon-like
peptide-1. Proc Natl Acad Sci USA 104, 15069–15074 (2007). Article ADS CAS PubMed Google Scholar * Kim, K. S., Egan, J. M. & Jang, H. J. Denatonium induces secretion of
glucagon-like peptide-1 through activation of bitter taste receptor pathways. Diabetologia 57, 2117–2125 (2014). Article CAS PubMed PubMed Central Google Scholar * Damak, S. et al.
Detection of sweet and umami taste in the absence of taste receptor T1r3. Science 301, 850–853 (2003). Article ADS CAS Google Scholar * Young, R. L. Sensing via intestinal sweet taste
pathways. Front Neurosci 5, 23 (2011). Article PubMed PubMed Central Google Scholar * Margolskee, R. F. Molecular mechanisms of bitter and sweet taste transduction. J Biol Chem 277, 1–4
(2002). Article CAS PubMed Google Scholar * Vandenbeuch, A. & Kinnamon, S. C. Why do taste cells generate action potentials? J Biol 8, 42 (2009). Article PubMed PubMed Central
Google Scholar * Zhang, Z., Zhao, Z., Margolskee, R. & Liman, E. The transduction channel TRPM5 is gated by intracellular calcium in taste cells. J Neurosci 27, 5777–5786 (2007).
Article CAS PubMed PubMed Central Google Scholar * Kim, K. S. et al. Transcriptomic analysis of the bitter taste receptor-mediated glucagon-like peptide-1 stimulation effect of quinine.
Biochip J 7, 386–392 (2013). Article CAS Google Scholar * Jiang, P. et al. Lactisole interacts with the transmembrane domains of human T1R3 to inhibit sweet taste. J Biol Chem 280,
15238–15246 (2005). Article CAS PubMed Google Scholar * Clapp, T. R. et al. Tonic activity of Galpha-gustducin regulates taste cell responsivity. FEBS Lett 582, 3783–3787 (2008). Article
CAS PubMed PubMed Central Google Scholar * Attele, A. S., Wu, J. A. & Yuan, C. S. Ginseng pharmacology: multiple constituents and multiple actions. Biochem Pharmacol 58, 1685–1693
(1999). Article CAS PubMed Google Scholar * Kinnamon, S. C. Taste receptor signalling - from tongues to lungs. Acta Physiol (Oxf) 204, 158–168 (2012). Article CAS Google Scholar *
Delghandi, M. P., Johannessen, M. & Moens, U. The cAMP signalling pathway activates CREB through PKA, p38 and MSK1 in NIH 3T3 cells. Cell Signal 17, 1343–1351 (2005). Article CAS
PubMed Google Scholar * Margolskee, R. F. et al. T1R3 and gustducin in gut sense sugars to regulate expression of Na+-glucose cotransporter 1. Proc Natl Acad Sci USA 104, 15075–15080
(2007). Article ADS CAS PubMed Google Scholar * Ma, J. et al. Effect of the artificial sweetener, sucralose, on gastric emptying and incretin hormone release in healthy subjects. Am J
Physiol Gastrointest Liver Physiol 296, G735–739 (2009). Article CAS PubMed PubMed Central Google Scholar * Steinert, R. E., Frey, F., Topfer, A., Drewe, J. & Beglinger, C. Effects
of carbohydrate sugars and artificial sweeteners on appetite and the secretion of gastrointestinal satiety peptides. Br J Nutr 105, 1320–1328 (2011). Article CAS PubMed Google Scholar *
Gerspach, A. C., Steinert, R. E., Schonenberger, L., Graber-Maier, A. & Beglinger, C. The role of the gut sweet taste receptor in regulating GLP-1, PYY and CCK release in humans. Am J
Physiol Endocrinol Metab 301, E317–325 (2011). Article CAS PubMed Google Scholar * Hwang, J. T. et al. Antiobesity effect of ginsenoside Rg3 involves the AMPK and PPAR-gamma signal
pathways. Phytother Res 23, 262–266 (2009). Article CAS PubMed Google Scholar * Kim, M. et al. The ginsenoside Rg3 has a stimulatory effect on insulin signaling in L6 myotubes. Biochem
Biophys Res Commun 389, 70–73 (2009). Article CAS PubMed Google Scholar * Kim, K. H. et al. Aqueous extracts of Anemarrhena asphodeloides stimulate glucagon-like pepetide-1 secretion in
enteroendocrine NCI-H716 cells. Biochip J 7, 188–193 (2013). Article CAS Google Scholar * Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402–408 (2001). Article CAS Google Scholar * Nelson, G. et al. Mammalian sweet taste receptors. Cell 106, 381–390 (2001).
Article CAS Google Scholar * Choi, E.-K. et al. Coix seed extract attenuates the high-fat induced mouse obesity via PPARγ and C/EBPα a downregulation. Mol Cell Toxicol 11, 213–221
(2015). Article CAS Google Scholar * Wong, G. T., Gannon, K. S. & Margolskee, R. F. Transduction of bitter and sweet taste by gustducin. Nature 381, 796–800 (1996). Article ADS CAS
PubMed Google Scholar Download references ACKNOWLEDGEMENTS The authors thank Dr. Robert F. Margolskee (Monell Chemical Senses Center, Philadelphia, PA, USA) for generously providing us
with a breeding pair of Gαgust−/− mouse. This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF), which is funded by the
Ministry of Education, Science and Technology (NRF-2013R1A1A2004960). AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Department of Biochemistry, College of Korean Medicine, Kyung Hee
University, 1 Heogi-dong, Dongdaemun-gu, Seoul, 130-701, Republic of Korea Ki-Suk Kim, Hea Jung Yang, In-Seung Lee, Kang-Hoon Kim, Jiyoung Park, Hyeon-Soo Jeong, Yoomi Kim & Hyeung-Jin
Jang * Department of Pathology, College of Korean Medicine, Kyung Hee University, 1 Heogi-dong, Dongdaemun-gu, Seoul, 130-701, Republic of Korea Kwang Seok Ahn * Western Seoul Center, Korea
Basic Science Institute, 150 Bugahyeon-ro, Seodaemun-gu, Seoul, 120-140, Republic of Korea Yoomi Kim & Yun-Cheol Na Authors * Ki-Suk Kim View author publications You can also search for
this author inPubMed Google Scholar * Hea Jung Yang View author publications You can also search for this author inPubMed Google Scholar * In-Seung Lee View author publications You can also
search for this author inPubMed Google Scholar * Kang-Hoon Kim View author publications You can also search for this author inPubMed Google Scholar * Jiyoung Park View author publications
You can also search for this author inPubMed Google Scholar * Hyeon-Soo Jeong View author publications You can also search for this author inPubMed Google Scholar * Yoomi Kim View author
publications You can also search for this author inPubMed Google Scholar * Kwang Seok Ahn View author publications You can also search for this author inPubMed Google Scholar * Yun-Cheol Na
View author publications You can also search for this author inPubMed Google Scholar * Hyeung-Jin Jang View author publications You can also search for this author inPubMed Google Scholar
CONTRIBUTIONS K.S.K. designed and performed GLP-1 assay, western blot analysis, qPCR and _in vivo_ studies using _db/db_ mice, analyzed the results and wrote the manuscript. H.J.Y. performed
cAMP assay, calcium imaging and _in vivo_ study using G_a_gust−/− mice. I.S.L., K.H.K., J.P. and H.S.J. participated in _in vivo_ studies using _db/db_ mice and G_a_gust−/− mice. K.S.A.
provided the ginsenosides. Y.K. and Y.C.N. performed and analyzed data of HPLC. H.J.J. designed the study and wrote the manuscript. ETHICS DECLARATIONS COMPETING INTERESTS The authors
declare no competing financial interests. ELECTRONIC SUPPLEMENTARY MATERIAL SUPPLEMENTARY INFORMATION RIGHTS AND PERMISSIONS This work is licensed under a Creative Commons Attribution 4.0
International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the
material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit
http://creativecommons.org/licenses/by/4.0/ Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Kim, KS., Jung Yang, H., Lee, IS. _et al._ The aglycone of ginsenoside Rg3 enables
glucagon-like peptide-1 secretion in enteroendocrine cells and alleviates hyperglycemia in type 2 diabetic mice. _Sci Rep_ 5, 18325 (2015). https://doi.org/10.1038/srep18325 Download
citation * Received: 16 July 2015 * Accepted: 16 November 2015 * Published: 17 December 2015 * DOI: https://doi.org/10.1038/srep18325 SHARE THIS ARTICLE Anyone you share the following link
with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt
content-sharing initiative