Common weeds as heavy metal bioindicators: a new approach in biomonitoring
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT Environmental pollution by heavy metals affects both urban and non-urban areas of Europe and the world. The use of bioindicator plants for the detection of these pollutants is a
common practice. An important property of potential bioindicators is their easy availability and wide distribution range, which means that they can be practically used over a wide area.
Therefore, common and widely distributed weeds: _Trifolium pratense_ L., _Rumex acetosa_ L., _Amaranthus retroflexus_ L., _Plantago lanceolata_ L., ornamental species _Alcea rosea_ L., and
_Lolium multiflorum_ L. var. Ponto were selected as a potential bioindicators of heavy metals (Cd, Pb, Cu, Zn). Plants were exposed in the same soil conditions in three sample sites in the
Poznań city. It was found that all species had heavy metal accumulation potential, especially _A. rosea_, _P. lanceolata_ and _L. multiflorum_ for Zn (BCF = 6.62; 5.17; 4.70) and _A. rosea,
P. lanceolata_ for Cd (BCF = 8.51; 6.94). Translocation of Cu and Zn was the most effective in _T. pratense_ (TFCu = 2.55; TFZn = 2.67) and in _A. retroflexus_ (TFCu = 1.50; TFZn = 2.23). Cd
translocation was the most efficient in _T. pratense_ (TFCd = 1.97), but PB was the most effective translocated in _A. retroflexus_ (TFPb = 3.09).. Based on physiological response to
stress, it was detected an increasing level of hydrogen peroxide (H2O2) in roots and leaves of all samples, with the highest in all organs of _A. rosea_. Enzymatic activity levels of CAT,
APOX, and also the marker of polyunsaturated fatty acid peroxidation MDA, were higher after 6 weeks of exposure in comparison to control samples and varied in time of exposure and between
species and exposure. After the experiment, in almost all samples we detected a reduction of chlorophyll content and relative water content, but in efficiency of photosynthesis parameters:
net photosynthesis rate, intercellular CO2 concentration and stomatal conductance, we noted increased values, which proved the relatively good condition of the plants. The examined weeds are
good bioindicators of heavy metal contamination, and their combined use makes it possible to comprehensively detection of environmental threats. SIMILAR CONTENT BEING VIEWED BY OTHERS LEVEL
OF HEAVY METALS AND ENVIRONMENTAL POLLUTION INDEX IN AHVAZ, SOUTHWEST IRAN Article Open access 26 June 2024 SOURCE ANALYSIS AND ECOLOGICAL RISK ASSESSMENT OF HEAVY METALS IN FARMLAND SOILS
AROUND HEAVY METAL INDUSTRY IN ANXIN COUNTY Article Open access 22 June 2022 HEAVY METAL SYNERGISTIC POLLUTION RISK ASSESSMENT IN THE SOIL-CROP SYSTEM OF THE NANYANG BASIN Article Open
access 06 June 2025 INTRODUCTION With the intensive development of human activities, urban areas have rapidly undergone significant and rapid changes. In urban areas, one of the most
important urban pollutants are metals and metalloids1, 2. Metals and metalloids are the subject of numerous studies because they are persistent and among the most widely disseminated
industrial pollutants3. The main sources of these elements are natural sources, such as natural weathering of crust, erosion, and anthropogenic activities, like urban runoff, agricultural
and industrial activities, and many others4. Exposure to heavy metals usually has subtle and chronic symptoms, moreover, exposure to airborne metals induces physiological responses in
organisms and broad health effects in humans5. Also, the contamination of dietary substances by heavy metals is known to have a range of adverse effects on humans, animals and plants6, 7. In
plants, their toxicity varies depending on the specific metal, plant spieces, pH, soil composition, and chemical form. Certain heavy metals are considered to be essential for development
and plant growth8. However, excess amounts of these elements can become toxic to plants9, thus affecting plants only negatively10. The exposure of plants to unfavorable environmental
conditions, including at higher concentrations of heavy metals, can cause an increase in the production of reactive oxygen species (ROS) such as singlet oxygen [(1) O2], superoxide
[(O2)−.)], hydrogen peroxide (H2O2), and hydroxyl radical (OH.). ROS modifies proteins, damages DNA and causes free radical oxidation of unsaturated fatty acids or other lipids the product
of which is MDA. The ROS detoxification process in plants is essential for the protection of plant cells, and therefore it seems that metal hyperaccumulating plants should have extremely
efficient antioxidative and detoxicative defense mechanisms, enabling growth and development in a polluted environment11. Plant responses and tolerance to heavy metal stress are dependent on
enzymatic antioxidants comprising ascorbate peroxidase (APOX), catalase (CAT), and the final product of polyunsaturated fatty acid peroxidation—malondialdehyde (MDA). These proteins take
part in ROS detoxification in plants12, and are present in practically all subcellular compartments. Usually, an organelle has more than one enzyme able to scavenge a single ROS13. As a
result of oxidative stress, photosynthetic processes are disturbed, from electron transport to carbon bonding. Limitation of any of these processes within the photosynthetic apparatus
reduces the ability of the chloroplast membrane to absorb light energy, increasing the ability to form oxidative radicals in the chloroplast, and as a consequence limits the productivity of
photosynthesis14. Identifying areas with higher concentrations of heavy metals, guidelines, and effective legislation are necessary. In addition, these metals should be subject to mandatory
monitoring due to their toxicity and possible bioaccumulation4. To control pollutants is a complex issue: the origin of pollutants and emission must be identified, critical emissions must be
controlled, techniques must be developed that are sufficiently sensitive and low-cost to allow simultaneous measurement of multiple contaminants, risks and economic factors must be
considered15. One inexpensive and simple method to determine the heavy metal concentration in the air and obtain information associated with the population’s exposure to air pollutants in a
particular ecosystem is biomonitoring16. Moreover, to obtain information about the changes in ecosystems, bioindicators can be used. Some plants are well known for their ability to
accumulate trace elements from the environment. Therefore, they have been used in a number of monitoring investigations, providing low-cost information regarding environmental quality with
the advantage of easy sampling. Various studies have used as bioindicators herbaceous plants (e.g., _Taraxacum officinale_, L., _Carduus nutans_ L., _Plantago major_ L., _Urtica dioica_ L.),
which are more common in urban environments (e.g.17,18,19,20). Previous studies have indicate the ability of selected species to bioaccumulate heavy metals. However, so far, they have not
been studied simultaneously, under the same contamination conditions, as well as taking into account the physiological response. The control of toxic elements contamination with the
simultaneous use of commonly available weeds like _Plantago lanceolata_ L.21, _Amaranthus retroflexus_ L.22, _Trifolium pratense_ L.1, _Rumex acetosa_23 and also known for the ability of
trace metals accumulation, old ornamental plant (_Alcea rosea_)24, seems to be a necessary procedure that enables comprehensive estimation of trace metals environment contamination. Selected
weeds have the basic characteristics of bioindicators, such as: long life cycle, wide geographic ranges, large numbers of occurrence and ease of determination. Our research focused on
checking whether selected weed species react in a characteristic way to changes in the environment (reaction to physical and chemical stress) depending on the place of occurrence. We made
these arrangements by using the active bioindication based on the exposure and observation of specific plant species. This is where the purpose of our research lies—to evaluate widespread
and very common bioindicators. Considering the above, the main objectives of this study were as follows: (i) to determine the accumulation level of trace metals (Cu, Zn, Cd and Pb) in
selected plant species exposed in even soil conditions in three research sites in the city; (ii) to assess the bioaccumulation potential of examined species; (iii) to establish translocation
of metals from soil to above-ground parts; (iv) to study the physiological conditions of plants; (v) to determine the activity of oxidative stress parameters; and (vi) to assess the
concentrations of enzymes of the antioxidative system. RESULTS The content of copper, zinc, cadmium and lead in the soil used for pot culture and in the tissues (roots and leaves) of all
samples of the studied species was examined, and then bioconcentration and translocation factors were calculated based on these results to assess bioaccumulation potential of examined
species. Moreover, the plant physiological responses for stress were detected for all samples of examined species. HEAVY METAL CONTENTS The heavy metal contents for all species at all
research sites showed the following tendency: Zn > Cu > Pb > Cd. This tendency was found for soil and plant organs (roots and leaves). In addition, for zinc and cadmium, the lowest
values were mostly observed in the soil, while for copper and lead their content was generally the highest in the soil, with only a few exceptions (Suppl. Table S1). Analyzing the data in
more detail, it was found that Cu, Zn, Cd and Pb concentrations in roots and leaves differ in all species. The highest Cu concentration in roots was found in _T. pratense_ (2C: 20.38 mg
kg−1); also a high value was recorded in _R. acetosa_ (3C: 10.51 mg kg−1) and in _L. multiflorum_ (1B: 8.30 mg kg−1). The highest Cu accumulation in leaves was detected in _R. acetosa_ (3C:
9.66 mg kg−1), in _T. pratense_ (2B: 9.20 mg kg−1) and in _A. rosea_ (4B: 8.13 mg kg−1). The highest Zn concentration in roots was detected in _L. multiflorum_ (1C: 81.13 mg kg−1), _P.
lanceolata_ (6C: 80.45 mg kg−1), _T. pratense_ (2C: 68.49 mg kg−1), _A. rosea_ (4B: 55.73 mg kg−1) and _A. retroflexus_ (5A: 52.62 mg kg−1). In _L. multiflorum_ leaves the highest Zn
concentration (1B: 172.45 mg kg−1) was noted; high Zn concentration in leaves of _A. rosea_ (4A: 135.85 mg kg−1) and _P. lanceolata_ (6C: 114.77 mg kg−1) was also found. In the soil samples
Zn concentration was lower than in plant tissues. The Cd amount varied in roots and leaves of studied species. In roots of _P. lanceolata_ (Control: 0.69 mg kg−1) we found the highest Cd
concentration; also high Cd concentration was found in _A. rosea_ (4A and 4B: 0.58 mg kg−1) roots. In leaves of _A. rosea_ (4B: 1.24 mg kg−1) we observed the highest Cd amount; also in _L.
multiflorum_ leaves (1C: 0.79 mg kg−1) and in _P. lanceolata_ (6A: 1.11 mg kg−1) high Cd concentration was detected in leaves. The highest Pb amount was found in _L. multiflorum_ roots (1C:
1.32 mg kg−1) as well as high Pb concentration in roots of _R. acetosa_ (3C: 0.75 mg kg−1). In leaf tissue of _L. multiflorum_ the highest Pb concentration was detected (1A: 1.21 mg kg−1),
in _R. acetosa_ Pb concentration in leaves reached 0.99 mg kg−1, and in _P. lanceolata_ it reached 0.77 mg kg−1. Pb concentration in soil samples was higher than in plants. However, two-way
ANOVA of species and site effect revealed significant influence (α ≤ 0.05) of both factors on all analyzed trace elements levels in roots and leaves. The both factors were found to have no
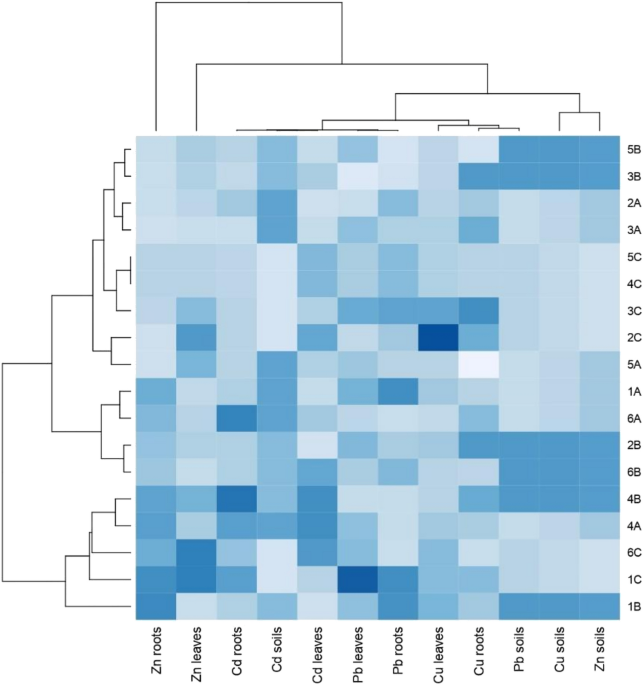
significant effect on the analyzed levels of these elements in soil, except of cadmium (one outlier observation in control) (Suppl. Table S2). Based on the cluster analysis with the
procedure for grouping objects and features (Fig. 1), taking into account all detected heavy metals, it can be found that there were differences between samples from sites A, B and C. The
soil most contaminated by Pb, Cu and Zn and Cd was from site B—the Botanical Garden. Two groups of samples were formed. The first consisted of 1B, 1C, 4A and 4B, and 6C. The second consisted
of four subgroups: the first subgroup included samples 6B, 2B, 6A and 1A; the second 5A, 2C; the third 3C, 4C, 5C; and the fourth 3A, 2A, 3B, 5B. Regarding the values of heavy metals in
roots of detected species, the highest concentration of Zn in roots was detected in _L. multiflorum_ (1B, 1C, 1A), _A. rosea_ (4A, 4B) and in _P. lanceolata_ (6A, 6C); the highest values of
Cd in roots was detected in _A. rosea_ (4A, 4B), _L. multiflorum_ (1C), and _P. lanceolata_ (6A) samples; the highest concentration of Pb in roots was noted in _L. multiflorum_ (1B, 1C, 1A),
_P. lanceolata_ (6B), _R. acetosa_ (3C), _A. rosea_ (4C), _A. retroflexus_ (5C) and in _T. pratense_ (2A). In sequence, taking into consideration Cu in roots, we noted the highest values in
_R. acetosa_ (3C, 3A and 3B), _T. pratense_ (2B, 2C), _A. rosea_ (4B) and _P. lanceolata_ (6A). Regarding the values of heavy metals in leaves of detected species, the highest
concentrations of Zn were noted in _L. multiflorum_ (1C), _P. lanceolata_ (6C and 3C), _A. rosea_ (4B), _A. retroflexus_ (5A), and _T. pratense_ (2C), while the highest amounts of Cd were
detected in leaves of the following samples: _P. lanceolata_ (6B, 6C), _A. rosea_ (4A, 4B, 4C), _T. pratense_ (2C), and _A. retroflexus_ (5C). Relatively high values of Pb in leaves were
detected in _L. multiflorum_ (1C, 1A), _T. pratense_ (2B), _R. acetosa_ (3C, 3A) and _A. retroflexus_ (5B), and finally, the highest concentrations of Cu in leaves were detected in the
following samples: _T. pratense_ (2C) and _R. acetosa_ in (3C). BIOCONCENTRATION AND TRANSLOCATION FACTOR The bioconcentration factors (BCF) exceeded a value of 1 for Zn and Cd in all plant
species. The highest values of Zn BCF were recorded in _L. multiflorum_ (1C: BCFZn = 6.62), in _A. rosea_ (4A: BCFZn = 5.17) and in _P. lanceolata_ (6C: BCFZn = 4.70); however, the highest
Cd BCF values were found in _A. rosea_ (4B: BCFCd = 8.51), in _P. lanceolata_ (6A: BCFCd = 6.94) and in 1C _L. multiflorum_ (BCFCd = 6.29). Cu and Pb bioconcentration was not as effective as
the first two elements, but it is worth mentioning that Cu bioconcentration factors of all detected samples exceed bioconcentration of Pb. Taking into account translocation of detected HMs,
the highest Cu TF value was detected in sample 2C of _T. pratense_ (TFCu = 2.55), in sample 6C of _P. lanceolata_ (TFCu = 1.55), in 5A sample _A. retroflexus_ (TFCu = 1.50), and in all
samples of _Lolium multiflorum_ (TFCu = 1.31–1.08). The Zn translocation factor was the highest in _T. pratense_ 2C (TFZn = 2.67), in all samples of _A. retroflexus_ (TFZn = 2.23–1.05), and
in _R. acetosa_ (TFZn = 1.28–1.11). The highest Cd translocation factor was detected in 2C _T. pratense_ (TFCd = 1.97), followed by _P. lanceolata_ (TFCd = 1.51–1.24), and in _R. acetosa_
(TFCd = 1.44–1.42). Regarding the Pb translocation factor, the highest values were detected in _A. retroflexus_ (TFPb = 3.09), in _P. lanceolata_ in 6B (TFPb = 2.25), and in sample 4A of _A.
rosea_ (TFPb = 1.90) (Table 1). PHYSIOLOGICAL CONDITION OF SPECIES After 6 weeks of the experiment, cell membrane stability (MSI) took values from 93.41% in _Alcea rosea_ sample 4C to the
highest values, more than 98%, in all _Plantago lanceolata_ samples, and in samples of _Trifolium pratense_, _Rumex acetosa_ and _Amaranthus retroflexus_. Dry mass content was highest
(23.11%) in _Amaranthus retroflexus_ sample 5A, and the lowest in _R. acetosa_ (8.01%) in sample 3C. In all species higher dry mass contents was detected in comparison to control samples.
RWC was the highest in _R. acetosa_ sample 3A (95.25%) and the lowest in _A. rosea_ sample 4C (62.53%). It should be noted that almost all samples were characterized by RWC above 90%.
Chlorophyll _a_ content ranged from 3.33 in _Amaranthus retroflexus_ to 11.02 in _Rumex acetosa_. Chlorophyll _b_ content ranged from 4.96 in _Trifolium pratense_ to 0.9 in _Amaranthus
retroflexus_. Chlorophyll _a_ + _b_ content ranged from 4.11 in _A. retroflexus_ to 15.36 in _R. acetosa._ Chlorophyll _a/b_ ratio ranged from 0.27 in _A. retroflexus_ to 0.58 in _T.
pratense_. Photosynthesis activity (_P_N) ranged in detected species from 7.24 in _Rumex acetosa_ to 28.28 in the _Trifolium pratense_ sample. Stomatal conductance (_g_s) ranged from 32.64
in _Amaranthus retroflexus_ to 188.86 in _Trifolium pratense_. _C_i intercellular CO2 concentration varies from 261.47 in _Amaranthus retroflexus_ to 528.20 in _Plantago lanceolata_ (Table
2). The graphical representation of the results by analysis of the first two principal components for heavy metal accumulation in leaves, roots and photosynthesis activity parameters in all
samples explained more than 47.81% of total variability (Fig. 2). A positive relationship was found between Zn-BCF, Cd-BCF in _L. multiflorum_ samples (1A, 1B, 1C), _T. pratense_ (2B)
samples and in _A. rosea_ (4B, 4C) samples. We also found a positive relationship between Pb TF and dry mass content in _P. lanceolata_ (6B). Another large group consists of correlated Zn
TF, Cu-TF, Cd TF with relative water content (RWC), cell membrane stability (MSI), chlorophyll b/a coefficient and net photosynthetic rate – _P_N. of _T. pratense_ (2A, 2C), _R. acetosa_
(3A, 3B) and _A. retroflexus_ (5C) samples. The last group was composed of Pb-BCF, Cu-BF correlated with chlorophyll content stomatal conductance (gs) and intercellular CO2 concentration
(_C_i) of _R. acetosa_ (3C) and _P. lanceolata_ (6A, 6C) samples. The profiles of changes and the level of hydrogen peroxide values in all species were similar in roots and in leaves, with
the highest amount of H2O2 ≈ 6 (nmol H2O2 × min−1 × mg protein−1) in roots of _R. acetosa_ and _A. rosea_ (4A). The highest amount of H2O2 ≈ 5 (nmol H2O2 × min−1 × mg protein−1) in leaves
was detected in all samples of _T. pratense_, _A. retroflexus_ and in _A. rosea_ sample 4A (Fig. S1). The profiles of changes and the level of CAT activity were similar in roots in leaves of
_L. multiflorum_, _R. acetosa_ and _A. retroflexus_. The highest activity of CAT ≈ 1.5 (nmol H2O2 × min−1 × mg protein−1) in roots and CAT ≈ 0.9 (nmol H2O2 × min−1 × mg protein−1) in leaves
was noted in _L. multiflorum_. It is worth noting that in _A. retroflexus_ and in _P. lanceolata_ high activity of CAT ≈ 0.9 (nmol H2O2 × min−1 × mg protein−1) was noted in roots. In leaves
the highest activity was noted in _L. multiflorum_ CAT ≈ 0.9 (nmol H2O2 × min−1 × mg protein−1) and in A. retroflexus CAT ≈ 0.55 (nmol H2O2 × min−1 × mg protein−1) (Fig. S2). APOX
activities were generally higher in roots than in leaves. In roots, the highest activity (APOX ≈ 0.065) was noted in _L. multiflorum_ and in _P. lanceolata_. In leaves APOX activity in all
samples was high, with the highest activity of APOX ≈ 0.03 in _L. multiflorum_ and in the _A. rosea_ sample (Fig. S3). The profiles of changes and the level of MDA activity were higher in
roots than in leaves of _T. pratense_, _R. acetosa_, _A. rosea_ and _A. retroflexus_. The highest level of MDA ≈ 12.0 in roots and in leaves (MDA ≈ 7.0) was detected in _A. rosea_ sample 4A
(Fig. S4). The graphical representation of the results by the analysis of the first two principal components for heavy metal accumulation in leaves, roots and photosynthesis activity
parameters in all samples explained more than 45% of the total variability (Fig. 3). A positive relationship was found between CAT activity in leaves and Cd and Cu in roots of _Plantago
lanceolata_ samples 6A and 6B and _Alcea rosea_ sample 4B. APOX activities in roots were related to Zn in roots in _Alcea rosea_ sample 4A. There was a positive relationship between hydrogen
peroxide amount and Cu and Zn in leaves of _Plantago lanceolata_ sample 6C. The next group consisted of APOX in leaves, MDA content in roots and the level of Pb in roots and leaves, the
level of Cu in leaves in _A. rosea_ 4C, _R. acetosa_ 3C, _A._ retroflexus 5C and _T. pratense_ 2C samples. Finally, there was a relationship between hydrogen peroxide in leaves, CAT activity
in roots in _A. retroflexus_ 5A and B, _R. acetosa_ 3A and _T. pratense_ 2A and B samples. The most intensive fluorescence DHE, indicating the presence of H2O2 in leaves, was observed in
the _T. pratense_ (2A), _Rumex acetosa_ (3A), A. rosea (4A) and _P. lanceolata_ (6A) samples (Fig. 4). DISCUSSION This comparative study of different weed species has shown their potential
for accumulation of HMs. _Lolium multiflorum_ var. Ponto was used as a known variety cumulating HMs19, 25, 26, but in our study it was found that this variety showed similar possibilities of
HM accumulation to the weed species selected for the experiment. Almost all the studied species showed significant potential for Zn and Cd accumulation compared to Cu and Pb. The efficiency
of accumulation was expressed as the BCF factor, and the metal displacement efficiency as the translocation factor (TF). For a hyperaccumulator plant, both of these factors should be
greater than unity27. Taking into account these two factors, the most effective bioindicator can be selected. Analyzing cadmium, which is the metal that is the most toxic to plants and
animals28, the highest content of Cd was found in the roots of _A. rosea_ and _P. lanceolata_. Very high bioconcentration coefficients calculated for these two species confirmed the
potential for this element to be concentrated in these species’ tissues. The usefulness of _A. rosea_ as a hyperaccumulator of Cd was noted previously by Liu et al.29 and Ubeynarayana et
al.30. Zn was also effectively accumulated mainly in leaves by _A. rosea_ and _P. lanceolata_. However, it is worth noting that Zn was well accumulated by all analyzed species. This ability
was also confirmed by bioconcentration factors determined for this species. Zinc as an essential micronutrient in plant nutrition is naturally taken up by plants, and the efficiency of
uptake depends on soil pH and phosphorus levels. In the countries of the European Union, soil is polluted with zinc, due to the use of sewage sludge for fertilization purposes or composts
made from them31. The amounts of Zn in unpolluted soils typically are lower than 125 ppm (125 mg kg−1) and in plants growing in these soils this metal concentration varies between 0.02 and
0.04 mg g−1 (20–40 mg kg−1) dry weight32. In our study, the highest concentration of Zn (≈ 29 mg/kg) was detected in Botanical Garden soil (B), which could be the consequence of use of
fertilizers in this area. However, according to Polish regulations on the permissible content of substances causing risk to human health and the environment, the obtained results do not
exceed the standards for soil at urban areas (Cu = 200 mg kg−1, Zn = 500 mg kg−1, Cd = 2 mg kg−1, Pb = 200 mg kg−1)33. Duan et al.34 noted that Zn was accumulated more in roots than in
leaves of _A. rosea_, which was confirmed in our study; the most effective accumulation of Zn was also detected in _A. rosea_ as well as in _P. lanceolata_ and _L. multiflorum_ var. Ponto.
We confirmed earlier information about mobility of Zn in plant tissues. The best Zn-TF transport efficiency was detected in _R. acetosa_. Longnecker et al.35 reported that in plants tolerant
of toxic levels of Zn, accumulation was observed in the root cortex and in their leaves. We detected that cadmium was effectively accumulated in the roots, but also efficiently transported
to the leaves of _A. rosea_, _P. lanceolata_, _T. pratense_ and _R. acetosa_ samples, as evidenced by high translocation factor (TF) values. If we take into account Zn and Cd together, we
should remember about the interaction between Zn and Cd, which consists in the mutual inhibition of the accumulation and sometimes translocation of elements in the plant36.Recent research
showed that the use of excess Zn, along with exposure to Cd under hydroponic conditions, mitigated Cd toxicity in plants by increasing all phenols and chlorophylls in the leaves, thereby
mitigating the adverse effects of Cd on photosynthetic function and oxygen secretion activity37. The authors suggested that these mechanisms are involved in Zn detoxification and protection
against Cd-induced structural and functional damage of the photosynthetic membranes. Pb accumulation was not so effective as mentioned earlier for Zn and Cd, but it is worth pointing out
that a relatively high concentration was found in leaves of all examined species, the highest in _L. multiflorum_ and _R. acetosa_. Barrutia et al.38 established that _R. acetosa_ had the
potential for bioconcentration of Pb (and also Zn and Cd), from highly contaminated mine soil. However, our research did not confirm the ability to hyperaccumulate this element, which could
be due to there being a relatively small amount in the soil of the studied sites. Lead was efficiently transported to the above part of the plant in selected samples of all examined species,
especially in _A. retroflexus_ and also in _A. rosea_ and _P. lanceolata_. Mobility of this ion in _Amaranthus spinosus_ was also confirmed by Yingping et al.39. In _Limbarda crithmoides_
and _Helianthus annuus_ Pb was not efficiently transported from roots to the leaves40 and it was only accumulated in roots. Taking into consideration Cu, we noted that _L. multiflorum_, _T.
pratense_, _R. acetosa_, _A. retroflexus_ and _P. lanceolata_ showed the ability to accumulate copper and its quick transport from the roots to the above-ground parts of the plant. Malizia
et al.1 confirmed that _T. pratense_ has the potential for copper bioconcentration. Our results also demonstrate a synergistic interaction between Cu, Cd and Zn during translocation of these
elements in shoots of _Trifolium pratense_ and _Amaranthus retroflexus_ (Fig. 2). Other researchers have also noted that _Amaranthus retroflexus_ accumulates metals such as Cd, Ni, Pb and
Cu in the aboveground parts41. In _Rumex acetosa_ an interaction between translocation of Cd and Zn (Fig. 2) was found, whereas other authors42 reported that Zn induced a decrease in Cd
uptake and a simultaneous increase in Zn accumulation in tomato plants. This suggested strong competition between Zn and Cd for the same membrane transporters. In the presence of heavy metal
ions, the plants in our study did not exhibit characteristic symptoms of their toxic effect on physiological activity. Dry mass, RWC, net photosynthetic rate _P_N, stomatal conductance gs
and intercellular CO2 concentration _C_i in plants after exposure were higher than in control species; only in one species—_R. acetosa—_did we detect a decrease in these parameters compared
to the control (Table 2). This may be due to relatively small amounts of heavy metals in the environment and relatively short exposure. The metal hyperaccumulating plants have an ability to
accumulate a relatively high level of HMs in their plant tissues, and they have developed a number of detoxification mechanisms for acclimation and tolerance of metals. The mechanism of Cd
tolerance has been extensively studied in many species43. Our research showed that species that accumulated Cd effectively triggered detoxification mechanisms to protect the function of the
photosynthetic apparatus against Cd stress44. Other authors observed a decrease in all photosynthesis parameters in _Amaranthus spinosus_39 and delayed chlorophyll fluorescence in _Lemna
minor_45. Contamination of plant tissues by heavy metals leads to the formation of ROS such as hydrogen peroxide (H2O2)12. We observed an increase in the level of ROS compared to control
plants in all plants after exposures. We observed higher levels of H2O2 in all plant organs of _T. pratense_, _R. acetosa_, _A. rosea_ and _P. lanceolata_. The increase in ROS production in
plants was associated with an increase in the activity of antioxidant enzymes. All examined species were characterized by the potential for accumulation of heavy metals. However, an
effective bioaccumulation process will depend on active detoxifying enzymes and the regulation of primary defense enzymes. We always observed the induction of antioxidant enzyme activity in
roots and leaves of plants, although there were no significant differences between the researched plants. Heavy metals modify membrane properties by interacting with functional groups of
membrane proteins and lipids. As a lipid peroxidation marker46, the measurement of malondialdehyde (MDA) content is used. In our research, MDA level activity after 2 weeks was higher than
after 6 weeks of exposure. The increase in MDA was induced by both essential metals such as Zn and non-essential metals such as Pb. It was detected that a higher MDA level in roots and
leaves in _Lolium multiflorum_, _Trifolium pratense_ and _Amaranthus retroflexus_ was correlated with high amounts of Pb in roots (Fig.). Lukatkin et al.22 reported that MDA levels in roots
and leaves of _Amaranthus retroflexus_ were correlated with high levels of Zn. Ascorbate peroxidase (APOX) isoforms play important and direct roles as protective elements against adverse
environmental conditions42. The decrease in membrane lipid peroxidation observed after 6 weeks may be due to activation of the ROS-inactivating antioxidant system. In our study APOX
activities were generally higher in leaves than in roots in all species. Activity of APOX was definitely lower than catalase, especially in the above-ground parts, which means that this
enzyme complements CAT catalytic activity. APOX may be responsible for controlling the levels of H2O2 as signal molecules, and the CAT function is to remove large amounts of oxygen during
oxidative stress12. The profiles of changes and the activity level of CAT were different between leaves and roots, but it is worth noting that in all sites with exposure of _A. retroflexus_
higher activity values of CAT were noted. Mohamed et al.47 showed in _Brassica juncea_ that the higher activity of antioxidant enzymes offers greater detoxification efficiency, which
provides better plant resistance against trace metal-induced oxidative stress. The results show the high accumulation potential of these species and their adequate physiological response to
stress. In order to detect environmental pollution, data from all tested species should be obtained, and such a procedure will more effectively determine the levels of risk for heavy metals
environment contamination. CONCLUSIONS Based on the obtained results, it can be concluded that all species showed varied but generally great potential for high accumulation of detected trace
elements. The concentrations of all elements in plant tissues were dependent on species, organ (root vs. shoot), and species-organ interactions. The physiological response of the studied
species to stress was correlated with the high content of the tested metals in the tissues. Plants exposed in different study sites showed different concentrations of trace elements in their
tissues. Due to the varying degree of the tested species ability to accumulate trace elements, in order to estimate the degree of environmental pollution by these compounds, we recommend
the simultaneous use of all species of weed which were tested in this work. In the future, finding a bioindicator among weeds that would have a whole set of excellent bioindicating features,
would provide a simple and cheap early warning system against the negative effects of changes in the ecosystem. MATERIALS AND METHODS In order to evaluate the bioaccumulation ability of Cd,
Pb, Cu and Zn by 6 selected plant species, an experiment was organized. The contents of these elements, their bioconcentration and translocation in plants and soil were also determined, as
well as the condition of the plants was evaluated. In addition, the results were analyzed using statistical analysis. MATERIALS Species for the study were selected due to their common, wide
range of occurrence, generally throughout Europe. For this investigation we selected species often found as weeds and as ornamental species in urbanized areas: _Lolium multiflorum_ L. (no.
1) is native to all Europe (except Finland), Western, Southern and Central Asia (except Uzbekistan), as well as Northern Africa. It was introduced to the Americas, South and East Africa,
Australia and East Asia (Hultén and Fries, 1986; POWO, 2019). For our purposes we used the _Lolium multiflorum_ variety Ponto, obtained from Norddeutsche Pflanzenzucht Hans-Georg Lembke KG
(Germany). It displayed phytoremediation potential for heavy metals48. _Trifolium pratense_ L. (no. 2) commonly known as red clover. It is a species native to: Europe, South, West and Middle
Asia as well as North-West Africa. Introduced and widespread in all continents except Antarctica49, 50. The seeds were collected by the authors from Wielkopolska rural areas; _Rumex
acetosa_ L. (no. 3), also known as common sorrel, is a herbaceous plant native to Europe, Asia, and North Africa (Morocco), and now it has spread to all continents except Australia and
Antarctica49, 50. The seeds were collected by the authors from Wielkopolska rural areas; _Alcea rosea_ L. (no. 4), the common hollyhock; it was imported into Europe from southwestern Asia as
an ornamental plant species before the fifteenth century. Since then, it is common ornamental plants in cities and widespread in the Americas, North Africa, and South Asia as a wilderness
species49, 50. In the present experiment we used _Alcea rosea_ L. The seeds were collected by the authors from Wielkopolska rural areas; _Amaranthus retroflexus_ L. (no. 5), the red-root
amaranth, is now found nearly worldwide. It is a species native to Mexico. It was introduced into all continents except Antarctica49, 50. The seeds were collected in the Poznań
agglomeration; _Plantago lanceolata_ L. (no. 6), ribwort plantain; this remarkably widespread species is native to all Europe, North Africa and West, South and Middle Asia but has been
introduced extremely widely elsewhere and now occurs e.g. in both Americas, Australia, New Zealand, Japan and in South and East Africa, where it thrives at high altitude49, 50. The seeds
were collected by the authors from Wielkopolska rural areas. We confirm that all methods including collection of plant material, were carried out in accordance with relevant guidelines and
regulations. ORGANIZATION OF THE EXPERIMENT The experiment was carried out during the growing season in 2021. The experiment started in April with planting the seeds in the control
greenhouse conditions (temperature 16–18 °C, no artificial light). 5 L pots with a standard mixture of peat and sand were used (pH 6.8, N: 230 mg L−1, P: 180 mg L−1, K: 350 mg L−1, Mg: 150
mg L−1). The content of the tested elements in potting soil at the beginning of the experiment was 4.151 ± 0.032 mg kg−1 for Cu, 15.03 ± 0.34 mg kg−1 for Zn 0.091 ± 0.004 mg kg−1 for Cd, and
4.302 ± 0.052 mg kg−1 for Pb. Seeds of each species were sown in individual pots in equal amounts. After germination, ten of the most vital and largest seedlings were left in the pot.
During germination and cultivation in the greenhouse, deionized water was used for plant irrigation. After 60 days, the plants were taken to exposure sites with various environmental
conditions. The three exposure sites were selected for these investigations, located in Poznań city. The first site (site A) was located in a residential area located on the right bank of
the Warta river (N: 52°23′53′′; E: 16°57′36′′), in the eastern part of the Poznań city (Fig. 5). High-density built-up areas (multi-family housing) dominated in the surrounding area of this
research site. The second exposure site (site B) was located in the Botanical Garden of the Adam Mickiewicz University in Poznań on the left bank of the Warta river (N: 52°25′14′′; E:
16°52′39′′), in the western part of the city. The immediate surroundings were green areas, whereas in the further surroundings of exposure site C there was a low-density built-up area
(single-family housing) and main road (N: 52°25′50′′; E: 16°54′58′′). At each site, three pots with plants of a given species were placed (in total, there were 18 pots per site). The
exposure of samples lasted 6 weeks (from June 1 to July 16, 2021). During the exposure, the plants were watered with distilled water and protected from the direct sun (shadowed) by naturally
occurring higher vegetation, without negative effect for air flow. Air pollution at research sites during exposition, were provided by the General Directorate for Environmental Protection
(Table S3). HEAVY METAL ANALYSIS PREPARATION SAMPLES In the laboratory, the plant samples were first purified with deionized water using Milli-Q Advantage A10 Water Purification Systems,
Merck Millipore (Merck, Darmstadt, Germany), and separated into leaves and roots. The soil samples from all pots were sieved (2 mm). To achieve constant dry weight, the plant and soil
samples were dried at 40 ± 3 °C in an electric oven (FD115, Binder, Germany). Digestion of the powdered samples of plant (homogenous samples from each pot) and soil were carried out in the
CEM Mars 5 Xpress microwave mineralization system (CEM, USA). From each plant, 0.3000 ± 0.0001 g of leaves or roots were placed in a Teflon vessel with 8 mL of concentrated (65%) HNO3
(analytical purity, Merck, Darmstadt, Germany) and 1 mL of H2O2 (Merck, Darmstadt, Germany). The program of digestion included the following stages—first stage: temperature to 80 °C, 10 min,
power 600 W; second stage: temperature 140 °C, 12 min, power 1200 W; third stage: temperature 185 °C, 15 min, power 1200 W. After the digestion steps using Qualitative Filter Papers (Grade
595: 4–7 μm Whatman, GB), the solutions were filtered, placed in flasks and made up to a final volume of 15 mL with deionized water. The analysis of the element's concentration in the
soil was performed in accordance with the PN-EN 16174 standards. Procedural blanks and reference materials were carried out in the same way as the samples in each digestion run. ANALYTICAL
PROCEDURE Elemental analysis of Cu, Zn, Cd and Pb was carried out using an inductively coupled plasma mass spectrometer (ICP-MS 7100 × Agilent, Santa Clara, CA, USA) equipped with an
octopole reaction system (ORS), MicroMist concentric nebulizer, quartz Scott double pass spray chamber, Ni cones, and a quadrupole mass spectrometer. The instrumental parameters were
optimized using the Tuning Solution (Agilent). The typical instrument operating conditions for ICP-MS spectrometers were as follows: 1550 W for RF power, 15 L min−1 for plasma gas flow rate,
0.98 L min−1 for nebulizer gas flow rate, 0.9 L min-1 for auxiliary gas flow rate. For the reduction of spectral interferences, helium mode was used. The non-spectral and matrix
interferences were reduced by diluting the samples and using an internal standard solution containing 10 µg L−1 Rh introduced in parallel with all analyzed solutions. High purity argon
(99.999%) was used as a nebulizer, auxiliary, and plasma gas for the ICP-MS (Messer, Chorzów, Poland). Calibration solutions were prepared by appropriate dilution of 10 mg L−1 of
multielemental stock solution in 5% HNO3 (Multi-Element Calibration Standard 3, PerkinElmer, MA, USA). The calibration curves were constructed in the concentration ranges: 0.05–50 µg L−1 for
Cd and Pb and 0.1–200.0 µg L−1 for Cu and Zn. QUALITY ASSURANCE To evaluate trueness and establish the traceability of the measurement result, certified reference materials (CRM) were used:
NIST SRM 1570a Trace Elements in Spinach Leaves (USA), NIST SRM 2711a Montana Soil. The validation parameters linearity, precision, limits of detection (LOD) and trueness were evaluated.
The linearity of the calibration curve was calculated as the correlation coefficient (R), the value of which is greater than 0.9996 for all analytes. The LOD for determined elements were
calculated according to LOD = 3.3 S/b, where S means standard deviation of the results obtained for the blank samples and b is the sensitivity (n = 5). The LOD values were as follows: Cd
0.007 µg g−1, Cu 0.036 µg g−1, Pb 0.008 µg g−1 and Zn 0.092 µg g−1. Precision values were calculated as the coefficient of variation (CV) (%) ranging from 0.8 to 2.3% for all elements.
Trueness was evaluated by applying the certified reference materials and expressed as recovery (%). Recovery values ranged from 97 to 102% for plants and from 93 to 98% for soil,
respectively. The results of Student’s t-test also confirmed that there were no significant differences between the measured concentration ± SD and the certified concentration ± standard
uncertainty. ACCUMULATION AND TRANSLOCATION FACTOR To estimate the efficiency of heavy metals’ phytoextraction by the studied plant species from three research sites, two factors were
calculated: bioconcentration and translocation. The ratio of heavy metal accumulation in root samples to heavy metal accumulation in soil samples was used to calculate bioconcentration
factor (BCF) of heavy metals51: $$BCF\, = \,HM \, concentration \, in \, roots \, \left( {{\text{mg kg}}^{ - 1} {\text{DW}}} \right)/ \, HM \, concentration \, in \, soil \, \left(
{{\text{mg kg}}^{ - 1} {\text{DW}}} \right).$$ Translocation factor (TF) is efficiency of the heavy metals’ transference to above-ground biomass52, with leaves and roots used in the
analysis: $$TF\, = \,HM \, accumulation \, in \, leaves \, \left( {{\text{mg kg}}^{ - 1} {\text{DW}}} \right)/ \, HM \, accumulation \, in \, roots \, \left( {{\text{mg kg}}^{ - 1}
{\text{DW}}} \right).$$ ANALYSIS OF PHYSIOLOGICAL CONDITIONS OF PLANTS DETERMINATION OF CHLOROPHYLL CONTENT To determine chlorophyll content, the experiment was performed in a laboratory,
where plants were brought from the three locations of the experiment. In order to avoid chlorophyll degradation, the experiment was carried out in subdued light during analyses and the
storage period. Three replicates for each plant sample were made. To determine the content of chlorophyll in plants, first undamaged leaves of the plant were cut and weighed approximately
0.100 g. The weighed samples were cut into smaller pieces and placed in a test tube, then 5 mL of 99.5% DMSO was added, the test tubes were closed with a stopper and they were placed in the
refrigerator for 24 h. After 24 h, samples were placed in a water bath at approximately 65 °C for 30–45 min to extract chlorophyll from the leaf blade. Then, the chlorophyll extract was
transferred to a 1 cm cuvette and the absorbance was measured on the Hach Lange DR-2800 spectrometer, at three wavelengths: 645 nm, 652 nm, and 663 nm. In parallel time, dry matter
determination was carried out for each plant sample. The chlorophyll content in samples was calculated using Arnon’s formula53. CELL MEMBRANE STABILITY (MSI) To determine cell membrane
stability from each plant, 2 cm2 of green leaves (without injury) were cut. Leaves which were cut were rinsed three times with double distilled water and were placed in 25 mL glass beakers,
immersed in 10 mL of distilled water then covered with aluminum foil and were put in a refrigerator for 24 h. The same process was repeated after 24 h: the distilled water was removed,
leaves were rinsed and were put in the same glass beakers, immersed in 10 mL of double distilled water, covered with aluminum foil and were put in a refrigerator for the next 24 h. After 24
h samples were taken out of the refrigerator and at room temperature their initial conductance was measured. After each measure samples were covered with aluminum foil. Then samples were
autoclaved at 0.5 atm, 105 °C for 30 min. After those processes samples were cooled at 25 °C and final conductance was measured. Their respective electric conductivities C1 and C2 were
measured by conductivity meters. The membrane stability index was calculated using the equation according to Almeselmani et al.54 formula. RELATIVE WATER CONTENT (RWC) Leaf relative water
content (RWC) estimation was done by cutting 3–4 pieces of leaf blade (without injuries). The pieces were weighed and placed in glass beakers. Leaves were pureed in 100 mL of distilled water
(completely submerged) and covered with aluminum foil, and they were placed in the refrigerator for 12 h. After 12 h, water was removed from the glass beakers and the samples were dried
with tissue paper and then were weighed again. Then after weighing samples, they were placed again in the same beakers and dried at 60 °C for 70 h. After 70 h, the samples were cooled in a
desiccator and were weighed again. The RWC value was calculated according to the formula by55. DRY MATTER CONTENT IN LEAVES The drying-weight method was used to determine dry matter content
in leaves. For each plant three replications were done. About 1 g of the plant's leaves were cut and were placed into a beaker, which were closed with a watch glass and placed in a
dryer for 24 h. Plants were dried at 105 °C until plants had a constant weight. The dry matter content was calculated based on the weight of the plants before and after drying, using formula
by Ostrowska et al.56. ANALYSIS OF ACTIVITY OF OXIDATIVE STRESS PARAMETERS AND LEVEL OF ENZYMES OF THE ANTIOXIDATIVE SYSTEM PHOTOSYNTHESIS At the beginning, in the middle, and at the end of
each exposure series we measured three intensity parameters: net photosynthesis (_P_N), intercellular CO2 concentration (_C_i,) stomatal conductance (_g_s). For measurement, matured leaves
were selected without mechanical injury. Gas exchange analysis was performed between 09:00 and 15:00 with the aid of the portable photosynthesis system Ci 340aa (CID Bioscience Inc., Camas,
WA, USA). To ensure similar conditions of measurements in the leaf chamber, stable conditions were provided: CO2 inflow concentration (410 µmol (CO2) mol−1), photosynthetic photon flux
density (PPFD) 1000 µmol (photon) m−2 s−1, a chamber temperature of 25 °C, and relative humidity of 50 ± 3%. HYDROGEN PEROXIDE CONTENT The hydrogen peroxide content was determined using the
method described by Patterson et al.57. The decrease in absorbance was measured at 508 nm using a UV–VIS spectrophotometer (Shimadzu Scientific Instruments, Japan). The reaction mixture
contained 50 mM phosphate buffer (pH 8.4) and reagents, 0.6 mM 4-(-2 pyridylazo) resorcinol, and 0.6 mM potassium-titanium oxalate (1:1). The corresponding concentration of H2O2 was
determined against the standard curve of H2O2. DETERMINATION OF ANTIOXIDATIVE ENZYME ACTIVITIES The activity of catalase (CAT, EC 1.11.1.6) was determined by directly measuring the
decomposition of H2O2 at 240 nm for 3 min as described by Aebi58 in a 50 mM phosphate buffer (pH 7.0) containing 5 mM H2O2 and enzyme extract. CAT activity was determined using the
extinction coefficient of 36 mM−1 cm−1 for H2O2. The activity of ascorbate peroxidase (APOX, EC 1.11.1.11) was assayed using the method described by Nakano and Asada59 by monitoring the rate
of ascorbate oxidation at 290 nm (extinction coefficient of 2.9 mM−1 cm−1) for 3 min. The reaction mixture consisted of 25–50 μL of supernatant, 50 mM phosphate buffer (pH 7.0), 20 μM H2O2,
0.2 mM ascorbate, and 0.2 mM EDTA. MEASUREMENT OF LIPID PEROXIDATION AND PROTEIN QUANTIFICATION Malondialdehyde (MDA) content was determined by reaction with thiobarbituric acid (TBA) as
described by Heath and Packer60. Total soluble protein contents were determined according to the method of Bradford61 using the Bio-Rad assay kit with bovine serum albumin as a calibration
standard. IN SITU DETECTION OF HYDROGEN PEROXIDE For the in vivo determination of hydrogen peroxide we used a modified version of the method described by Afzal et al.62. All plant specimens
were submerged for 12 h in 4 µM dichlorodihydrofluorescein diacetate (DCFH-DA) in 5 mM dimethyl sulfoxide (DMSO). After rinsing with 50 mM phosphate buffer (pH 7.4), the roots were observed
with a confocal microscope (Zeiss LSM 510, Axiovert 200 M, Jena, Germany) equipped with no. 10 filter sets (excitation 450–490 nm, emission 520 nm or more). STATISTICAL ANALYSIS Descriptive
statistical analysis was performed to assess the concentrations of heavy metals in examined plant species from different samples and also concentration of defense system and physiological
parameters. Statistical analysis was performed for 72 pots (separately for leaves, roots and soil). All samples followed assumptions of distribution normality and homogeneity. Analysis of
variance (two-way ANOVA) was used to assess the significance of differences between species and location for all parameters, and finally, the Scheffé test was applied to show the existence
of uniform groups of objects (soils, roots and leaves, separately) (α ≤ 0.05). Principal component analysis (PCA) was performed to evaluate associations between elemental contents and
different cities and determine interactions between independent variables (relations between elemental contents in species, location and physiological parameters), without any a priori
assumptions. Cluster analyses with procedure grouping objects and features, were performed using R platform (R Core 2014), to find similarities between sites, species, and heavy metal
accumulations. Data were visualized using heat maps to compare the concentration of a particular group of elements in plants and soils at specific research sites, with two-dimensional
variables (research sites, element) represented by colors. Statistical analyses were carried out using statistical software (Statistica 13.1) and R computer platform (R Core, 2014). DATA
AVAILABILITY All data included in this study are available upon request by contact with the corresponding author. REFERENCES * Malizia, D., Giuliano, A., Ortaggi, G. & Masotti, A. Common
plants as alternative analytical tools to monitor heavy metals in soil. _Chem. Cent. J._ 6(2), 1–10 (2012). Google Scholar * Koller, M., & Saleh, H. M. Introducing heavy metals. in
_Heavy Metals_ (eds. El-Din, H., Saleh, M. & Aglan, R. F.). https://doi.org/10.5772/intechopen.74783 (Intech Open, 2018). * Carreras, H. A. & Pignata, M. L. Biomonitoring of heavy
metals and air quality in Cordoba City, Argentina, using transplanted lichens. _Environ. Pollut._ 117(1), 77–87 (2002). Article CAS PubMed Google Scholar * Morais, S., Costa, F. G. &
Lourdes Pereira, M. Heavy metals and human health. in _Environmental Health—Emerging Issues and Practice_ (eds. El-Din, H., Saleh, M. & Aglan, R. F.).
https://doi.org/10.5772/intechopen.71185 (IntechOpen, 2012). * Mohanraj, R. P. A. & Priscilla, A. T. Heavy metals in airborne particulate matter of urban Coimbatore. _Arch. Environ.
Contam. Toxicol._ 47, 162–167 (2004). Article CAS PubMed Google Scholar * Nagajyoti, P. C., Lee, K. D. & Sreekanth, T. V. M. Heavy metals, occurrence and toxicity for plants: A
review. _Environ. Chem. Lett._ 8, 199–216 (2010). Article CAS Google Scholar * Ackova, D. G. Heavy metals and their general toxicity for plants. _Plant Sci. Today_ 5, 14–18 (2018).
Article CAS Google Scholar * Mukesh, K. R., Kumar, P., Singh, M. & Singh, A. Toxic effect of heavy metals in livestock health. _Veterin World_ 1, 28–30 (2008). Google Scholar *
Djingova, R. & Kuleff, I. Instrumental techniques for trace analysis. in _Trace Elements: Their Distribution and Effects in the Environment_ (Vernet, J.P. Ed.). Vol. 4. 137–185
(Elsevier, 2000). * Asati, A., Pichhode, M. & Nikhil, K. Effect of heavy metals on plants: An overview. _Int. J. Appl. Innov. Eng. Manag. (IJAIEM)_ 5(5–3), 2319–4847 (2016). Google
Scholar * Małecka, A., Ciszewska, L., Staszak, A. & Ratajczak, E. Relationship between mitochondrial changes and seed aging as a limitation of viability for the storage of beech seed
(_Fagus sylvatica_ L.). _PeerJ_ https://doi.org/10.7717/peerj.10569 (2021). Article PubMed PubMed Central Google Scholar * Małecka, A., Konkolewska, A., Hanć, A., Kmita, H. &
Jarmuszkiewicz, W. Insight into the phytoremediation capability of _Brassica juncea_ (v. Malopolska): Metal accumulation and antioxidant enzyme activity. _Int. J. Mol. Sci._ 20(18), 4355
(2019). Article PubMed PubMed Central Google Scholar * Scandalios, J. G. Oxidative stress: Molecular perception and transduction of signal triggering antioxidant gene defenses. _Braz. J.
Med. Biol. Res._ 38, 995–1014 (2005). Article CAS PubMed Google Scholar * Moustakas, M. _et al._ Spatial heterogeneity of cadmium effects on salvia sclarea leaves revealed by
chlorophyll fluorescence imaging analysis and laser ablation inductively coupled plasma mass spectrometry. _Materials._ https://doi.org/10.3390/ma12182953 (2019). Article PubMed PubMed
Central Google Scholar * Rucandio, M. I., Petit-Domínguez, M. D., Fidalgo-Hijano, C. & García-Giménez, R. Biomonitoring of chemical elements in an urban environment using arboreal and
bush plant species. _Environ. Sci. Pollut. Res._ 18(1), 51–63 (2011). Article CAS Google Scholar * Giampaoli, P., Wannaz, E., Tavares, A. & Domingos, M. Suitability of _Tillandsia
usneoides_ and _Aechmea fasciata_ for biomonitoring toxic elements under tropical seasonal climate. _Chemiosphere_ https://doi.org/10.1016/j.chemosphere.2016.01.080 (2016). Article Google
Scholar * Diatta, J., Grzebisz, W. & Apolinarska, K. A study of soil pollution by heavy metals in the city of Poznan (Poland) using dandelion (_Taraxacum officinale_ Web) as a
bioindicator. _Electron. J. Pol. Agric. Univ._ 6(2), 638 (2003). Google Scholar * Radulescu, C. _et al._ Heavy metal accumulation and translocation in different parts of _Brassica oleracea_
L. _Rom. J. Phys._ 58(9–10), 1337–1354 (2013). CAS Google Scholar * Borowiak, K. _et al._ Relations between photosynthetic pigments macro-elements contents and selected trace elements
accumulated in _Lolium multiflorum_ L. exposed to ambient air conditions. _Acta Biol. Cracoviensia Ser. Bot._ 60(1), 35–44 (2018). CAS Google Scholar * Lisiak-Zielińska, M. _et al._ How
polluted are cities in central Europe?—Heavy metal contamination in _Taraxacum officinale_ and soils collected from different land use areas of three representative cities. _Chemosphere_
266, 129113 (2021). Article ADS PubMed Google Scholar * Nagórska-Socha, A., Ptasiński, B. & Kita, A. Heavy metal bioaccumulation and antioxidative responses in _Cardaminopsis
arenosa_ and _Plantago lanceolata_ leaves from metalliferous and non-metalliferous sites: A field study. _Ecotoxicology_ 22, 1422–1434. https://doi.org/10.1007/s10646-013-1129-y (2013).
Article CAS Google Scholar * Lukatkin, A. S., Dmitry, I., Bashmakov, D. I., Harbawee, W. E. Q. & Teixeira da Silva, J. A. Assessment of physiological and biochemical responses of
_Amaranthus retroflexus_ seedlings to the accumulation of heavy metals with regards to phytoremediation potential. _Int. J. Phytoremediat._ 23(3), 219–230 (2021). Article CAS Google
Scholar * Ligarda-Samanez, C. A. _et al._ Modified polymeric biosorbents from _Rumex acetosella_ for the removal of heavy metals in wastewater. _Polymers_ 14(11), 2191.
https://doi.org/10.3390/polym14112191 (2022). Article CAS PubMed PubMed Central Google Scholar * Kaya, I. & Gülser, F. Original research determining heavy metal contents of
Hollyhock (_Alcea rosea_ L.) in roadside soils of a Turkish Lake Basin. _Pol. J. Environ. Stud._ 27(5), 2081–2087. https://doi.org/10.15244/pjoes/79270 (2018). Article CAS Google Scholar
* Klumpp, A. _et al._ Airborne trace element pollution in 11 European cities assessed by exposure of standardised ryegrass cultures. _Atmos. Environ._ 43, 329–333 (2009). Article ADS CAS
Google Scholar * Maciejewska-Malina, J. & Maciejewska, A. Uptake of heavy metals by darnel multifloral (_Lolium multiflorum_ Lam.) at diverse soil reaction and organic matter content.
_Soil Sci. Annu._ 64(1), 19–22 (2013). Article Google Scholar * Yanqun, Z. _et al._ Hyperaccumulation of Pb, Zn and Cd in herbaceous grown on lead–zinc mining area in Yunnan, China.
_Environ. Int._ 31(5), 755–762 (2005). Article PubMed Google Scholar * Kabata-Pendias, A. & Pendias, H. _Trace Elements in Soils and Plants_ 3rd edn. (CRC Press, 2001). Google Scholar
* Liu, J., Zhou, Q., Song, W. & Ting, S. Cadmium tolerance and accumulation of _Althaea rosea_ Cav. and its potential as a hyperaccumulator under chemical enhancement. _Environ. Monit.
Assess._ 149, 419–427 (2008). Article PubMed Google Scholar * Ubeynarayana, N., Paramsothy, J., Bishop, P., RobertoCalveloPereira, R. & Anderson, C. W. N. Effect of soil cadmium on
root organic acid secretion by forage crops. _Environ. Pollut._ https://doi.org/10.1016/j.envpol.2020.115839 (2021). Article PubMed Google Scholar * Barajas-Aceves, M. Comparison of
different microbial biomass and activity measurement methods in metal-contaminated soils. _Biores. Techn._ 96, 1405–1414 (2005). Article CAS Google Scholar * Tsonev, T. & Cebola
Lidon, F. J. C. Zinc in plants—An overview. _Emir. J. Food Agric._ 24(4), 322–333 (2012). Google Scholar * Journal of Laws of the Republic of Poland item 1395. _Regulation of the Minister
of the Environment (September 1, 2016) on the Method of Assessing the Pollution of the Earth’s Surface_ (2016). * Duan, Y., Zhang, Y. & Zhao, B. Lead, zinc tolerance mechanism and
phytoremediation potential of _Alcea rosea_ (Linn.) Cavan and _Hydrangea macrophylla_ (Thunb.) Ser. and ethylenediaminetetraacetic acid effect. _Environ. Sci. Pollut. Res. Int._ 29(27),
41329–41343 (2022). Article CAS PubMed Google Scholar * Longnecker, N. E. & Robson, A. D. Distribution and transport of zinc in plants. in _Zinc in Soils and Plants. Developments in
Plant and Soil Sciences_ (ed. Robson, A. D.). Vol. 55. (Springer, 1993). * Du, J. _et al._ The presence of zinc reduced cadmium uptake and translocation in Cosmos bipinnatus seedlings under
cadmium/zinc combined stress. _Plant Physiol. Biochem._ 151, 223–232 (2020). Article CAS PubMed Google Scholar * Dobrikova, A. _et al._ Combined impact of excess zinc and cadmium on
elemental uptake, leaf anatomy and pigments, antioxidant capacity, and function of photosynthetic apparatus in Clary Sage (_Salvia sclarea_ L.). _Plants_ 11, 2407 (2022). Article CAS
PubMed PubMed Central Google Scholar * Barrutia, O., Epelde, J. I., García-Plazaola, C., Garbisu, J. & Becerri, M. Phytoextraction potential of two _Rumex acetosa_ L. accessions
collected from metalliferous and non-metalliferous sites: Effect of fertilization. _Chemosphere_ 74(2), 259–264 (2009). Article ADS CAS PubMed Google Scholar * Yingping, H. _et al._
Effects of lead and cadmium on photosynthesis in _Amaranthus spinosus_ and assessment of phytoremediation potential. _Int. J. Phytorem._ 21(10), 1041–1049 (2019). Article Google Scholar *
Dridi, N., Bouslimi, H., Caçador, I. & Sleimi, N. Lead tolerance, accumulation and translocation in two Asteraceae plants: _Limbarda crithmoides_ and _Helianthus annuus_. _S. Afr. J.
Bot._ 150, 986–996 (2022). Article CAS Google Scholar * Cherif, J., Mediouni, C., Ammar, W. B. & Jemal, F. Interactions of zinc and cadmium toxicity in their efects on growth and in
antioxidative systems in tomato plants _(Solanum lycopersicum_). _J. Environ. Sci._ 23, 837–844 (2011). Article CAS Google Scholar * Bayçu, G. _et al._ Cadmium-zinc accumulation and
photosystem II responses of _Noccaea caerulescens_ to Cd and Zn exposure. _Environ. Sci. Pollut. Res._ 24, 2840–2850 (2017). Article Google Scholar * Dobrikova, A. G. _et al._ Cadmium
toxicity in _Salvia sclarea_ L: An integrative response of element uptake, oxidative stress markers, leaf structure and photosynthesis. _Ecotoxicol. Environ. Saf._ 209, 111851 (2021).
Article CAS PubMed Google Scholar * Drinovec, L., Drobne, D., Jerman, I. & Zrimec, A. Delayed fluorescence of _Lemna minor_: A biomarker of the effects of copper, cadmium, and zinc.
_Bull. Environ. Contam. Toxicol._ 72, 896–902 (2004). Article CAS PubMed Google Scholar * Morales, M. & Munné-Bosch, S. Malondialdehyde: Facts and artifacts. _Plant Physiol._ 180(3),
1246–1250 (2019). Article CAS PubMed PubMed Central Google Scholar * Caverzan, A. _et al._ Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection.
_Genet. Mol. Biol._ 35(4), 1011–1019 (2012). Article CAS PubMed PubMed Central Google Scholar * Mohamed, A. A., Castagna, A., Ranieri, A. & di Toppi, L. S. Cadmium tolerance in
_Brassica juncea_ roots and shoots is affected by antioxidant status and phytochelatin biosynthesis. _Plant Physiol. Biochem._ 57, 15–22 (2012). Article CAS PubMed Google Scholar * Cui,
E., Cui, B., Fan, X., Li, S. & Feng, G. Ryegrass (_Lolium multiflorum_ L.) and Indian mustard (_Brassica juncea_ L.) intercropping can improve the phytoremediation of antibiotics and
antibiotic resistance genes but not heavy metals. _Sci. Total Environ._ 25(784), 147093 (2021). Article ADS Google Scholar * Hultén, E. & Fries, M. _Atlas of North European Vascular
Plants. North of the Tropic of Cancern. Introduction to Taxonomic index to the maps 1-996. Maps 1-996I_. (Koeltz Scientific Books, 1986). * POWO.
https://www.powo.science.kew.org/taxon/urn:lsid:ipni.org:names:18423-1 (2019). * Ali, H., Khan, E. & Sajad, M. A. Phytoremediation of heavy metals—Concepts and applications.
_Chemosphere_ 91(7), 869–881 (2013). Article ADS CAS PubMed Google Scholar * Rezvani, M. & Faezeh, Z. Bioaccumulation and translocation factors of cadmium and lead in _Aeluropus
littoralis_. _Aust. J. Agric. Eng._ 2(4), 114–119 (2011). Google Scholar * Arnon, D. & Copper, I. Enzymes in isolated chloroplasts. Polyphenoloxidase in _Beta vulgaris_. _Plant
Physiol._ 24, 1–15 (1949). Article CAS PubMed PubMed Central Google Scholar * Almeselmani, M. _et al._ Physiological traits associated with drought tolerance of syrian durum wheat
varieties under rainfed condition. _Indian J. Plant Physiol._ 17(2), 166–169 (2012). Google Scholar * Creus, C. M., Sueldo, R. J. & Barass, C. Water relations and yield in
_Azospirillum_ inoculated wheat exposed to drought in the field. _Can. J. Bot._ 82(2), 273–281 (2004). Article Google Scholar * Ostrowska-Gumkowska, B. & Ostrowska-Czubenko, J. Effect
of comonomer content on thermal degradation of anionically modified poly(ethylene terephthalate). _Eur. Polymer J._ 27(7), 681–686 (1991). Article CAS Google Scholar * Patterson, B. D.,
Macrae, E. A. & Ferguson, I. B. Estimation of hydrogen peroxide in plant extracts using titanium(IV). _Anal. Biochem._ 139, 487–492 (1984). Article CAS PubMed Google Scholar * Aebi,
H. & Catalase, E. _Methods of Enzymatic Analysis _(ed. Bergmeyer, H. U.)_. _273–286 (Chemie, 1983). * Nakano, Y. & Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific
peroxidase in spinach chloroplasts. _Plant Cell Physiol._ 22, 867–880 (1981). CAS Google Scholar * Heath, R. L. & Packer, L. Photoperoxidation in isolated chloroplasts: Kinetics and
stoichiometry of fatty acid peroxidation. _Arch. Biochem. Biophys._ 125, 189–198 (1968). Article CAS PubMed Google Scholar * Bradford, M. M. A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. _Anal. Biochem._ 72(1–2), 248–254 (1976). Article CAS PubMed Google Scholar * Afzal, M.
_et al._ Method to overcome photoreaction, a serious drawback to the use of dichlorofluorescin in evaluation of reactive oxygen species. _Biochem. Biophys. Res. Commun._ 304, 619–624 (2003).
Article CAS PubMed Google Scholar Download references ACKNOWLEDGEMENTS We are grateful to Barbara Andrzejewska for technical support in the laboratory. Our sincere thanks are due to
Erjon Qorri for help in graphical preparation of statistical results. We also thank Prof. Zbigniew Celka from the Department of Biology Adam Mickiewicz University in Poznań for assistance in
collecting seeds. We are indebted to Prof. Justyna Wiland-Szymańska, who allowed us to use experimental plots in the Botanical Garden of the Adam Mickiewicz University in Poznań. Our
sincere thanks are due to Norddeutsche Pflanzenzucht Hans-Georg Lembke KG (Germany) for sending the seed material of the _Lolium multiflorum_ variety Ponto. FUNDING This work was financially
support by Faculty of Environmental and Mechanical Engineering Poznań University of Life Sciences. AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Department of Ecology and Environmental
Protection, Faculty of Environmental and Mechanical Engineering, Poznań University of Life Sciences, Piątkowska 94C, 60-649, Poznań, Poland A. Cakaj, M. Lisiak-Zielińska, K. Borowiak &
M. Drapikowska * Department of Trace Analysis, Faculty of Chemistry, Adam Mickiewicz University, Uniwersytetu Poznańskiego 8, 61-614, Poznań, Poland A. Hanć * The Cancer Epidemiology and
Prevention Unit, Greater Poland Cancer Centre, Garbary 15 Street, 61-866, Poznan, Poland A. Małecka Authors * A. Cakaj View author publications You can also search for this author inPubMed
Google Scholar * M. Lisiak-Zielińska View author publications You can also search for this author inPubMed Google Scholar * A. Hanć View author publications You can also search for this
author inPubMed Google Scholar * A. Małecka View author publications You can also search for this author inPubMed Google Scholar * K. Borowiak View author publications You can also search
for this author inPubMed Google Scholar * M. Drapikowska View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS A.C.-Conceptualization, field
research, biochemical analysis, partly draft preparation. M.L.Z.-Conceptualization, biochemical analysis, partly draft preparation, edition. A.H.-Chemical analysis, partly draft preparation.
A.M.-Biochemical analysis, confocal microscopy analysis. K.B.-Biochemical analysis, partly draft preparation. M.D.-Conceptualization, statistics evaluation, partly draft preparation and
edition. CORRESPONDING AUTHOR Correspondence to M. Drapikowska. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION PUBLISHER'S
NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION. RIGHTS AND
PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any
medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The
images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is
not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission
directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Cakaj,
A., Lisiak-Zielińska, M., Hanć, A. _et al._ Common weeds as heavy metal bioindicators: a new approach in biomonitoring. _Sci Rep_ 13, 6926 (2023). https://doi.org/10.1038/s41598-023-34019-9
Download citation * Received: 27 February 2023 * Accepted: 22 April 2023 * Published: 28 April 2023 * DOI: https://doi.org/10.1038/s41598-023-34019-9 SHARE THIS ARTICLE Anyone you share the
following link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer
Nature SharedIt content-sharing initiative