Selection of rna-based evaluation methods for tumor microenvironment by comparing with histochemical and flow cytometric analyses in gastric cancer
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT Understanding the tumor microenvironment (TME) and anti-tumor immune responses in gastric cancer are required for precision immune-oncology. Taking advantage of next-generation
sequencing technology, the feasibility and reliability of transcriptome-based TME analysis were investigated. TME of 30 surgically resected gastric cancer tissues was analyzed by RNA-Seq,
immunohistochemistry (IHC) and flow cytometry (FCM). RNA-Seq of bulk gastric cancer tissues was computationally analyzed to evaluate TME. Computationally analyzed immune cell composition was
validated by comparison with cell densities established by IHC and FCM from the same tumor tissue. Immune cell infiltration and cellular function were also validated with IHC and FCM. Cell
proliferation and cell death in the tumor as assessed by RNA-Seq and IHC were compared. Computational tools and gene set analysis for quantifying CD8+ T cells, regulatory T cells and B
cells, T cell infiltration and functional status, and cell proliferation and cell death status yielded an excellent correlation with IHC and FCM data. Using these validated
transcriptome-based analyses, the immunological status of gastric cancer could be classified into immune-rich and immune-poor subtypes. Transcriptome-based TME analysis is feasible and is
valuable for further understanding the immunological status of gastric cancer. SIMILAR CONTENT BEING VIEWED BY OTHERS POTENTIALITY OF MULTIPLE MODALITIES FOR SINGLE-CELL ANALYSES TO EVALUATE
THE TUMOR MICROENVIRONMENT IN CLINICAL SPECIMENS Article Open access 11 January 2021 TUTORIAL: INTEGRATIVE COMPUTATIONAL ANALYSIS OF BULK RNA-SEQUENCING DATA TO CHARACTERIZE TUMOR IMMUNITY
USING RIMA Article 30 June 2023 MULTI-OMICS FEATURES OF IMMUNOGENIC CELL DEATH IN GASTRIC CANCER IDENTIFIED BY COMBINING SINGLE-CELL SEQUENCING ANALYSIS AND MACHINE LEARNING Article Open
access 18 September 2024 INTRODUCTION The prognosis of locally advanced and metastatic gastric cancer remains poor, resulting in metastatic gastric cancers now being the fourth leading cause
of cancer death globally1. Immune checkpoint inhibitors (ICIs) targeting the PD-1/PD-L1 pathway were approved for metastatic gastric cancer in 2017; this has improved the prognosis, but the
benefits are nonetheless limited2,3. However, clinical indications for ICI are expanding, and different treatment strategies such as earlier administration or combinations with other
chemotherapies are now becoming feasible4,5. To optimize these approaches, a rational design of combination immunotherapies for gastric cancer requires a better understanding of the tumor
microenvironment (TME) and anti-tumor immune responses, which in turn requires optimal analytical methods for use in the clinic. The Cancer Genome Atlas Research Network (TCGA) advocates
comprehensive molecular characterization of gastric cancer based on RNA sequencing from bulk tissue (bulk RNA-Seq)6. Here, we undertook bulk RNA-Seq of gastric cancer and evaluated the TME
based on the "Cancer-Immunity Cycle" that dynamically represents the intratumoral immune response7. We then proposed an immunogram classification for gastric cancer8. Although good
therapeutic responsiveness to ICI has been shown in MSI-type and EBV-type gastric cancer9, responses to ICI in other types are poor. An immunosuppressive TME with complex heterogeneity
might be one important reason for this difficulty in treating gastric cancer and remains to be further investigated. RNA-Seq is an attractive tool for the interrogation of the transcriptome
of a tumor and its microenvironment. It is possible to perform RNA-Seq analysis from either fresh-frozen or fresh tissue, with even a tiny piece of biopsy specimen sufficing. Compared to
immunohistochemistry (IHC) and flow cytometry (FCM), many aspects of the immune response in the tumor can be investigated simultaneously with a very large number of markers by single method
RNA-Seq. Although RNA-Seq provides comprehensive transcriptomic data, extracting biological insight and deducing the presence of different immune cells from such data requires computational
analytical methods. However, there is a recognized problem regarding potential discrepancies between the quantification of cell populations based on transcriptomic data and the actual amount
of the corresponding cell types estimated by IHC or FCM10. Therefore, further efforts to refine and improve the analysis of transcriptomic signatures are needed for their clinical
application. In this study, we examined the correlation between transcriptome-based analysis and IHC or FCM assessments, currently considered the gold standard for quantifying cell type
composition and functional status. We selected the appropriate gene sets and computational analysis frameworks that reflect the TME of gastric cancer in terms of tumor-infiltrating
lymphocytes, proliferation of tumor cells and immune cells, and tumor cell death. Using these validated computational methods, we propose a novel RNA-based evaluation of intratumoral immune
responses in gastric cancer. RESULTS PATIENTS’ CHARACTERISTICS The characteristics of patients in the BKT cohort are shown in Table 1 and Supplementary Table S1. Five of 30 patients had
Stage IV disease and underwent palliative surgery. The HER2-positive rate was 23.3%, and the percentage of each TCGA molecular classification was as follows: MSI (16.7%), EBV (10.0%), GS
(13.3%), and CIN (60.0%). Thus, the cohort appears to be representative, with no significant deviations from previous reports6,11. EVALUATION OF TRANSCRIPTOME-BASED CELL TYPE QUANTIFICATION
The different transcriptome-based immune cell quantification methods listed in Table 2 were applied to RNA-Seq data of 30 gastric cancer patients (Supplementary Table S2). Multiple immune
cells were computationally quantified from one RNA-Seq dataset. These methods can be divided into two categories, namely, values for the expression of marker genes (marker-based approach) or
the deconvolution approach. The output scores are the absolute value of each cell population or the fraction (%) of the total cells. Therefore, certain defined cell type data can be
compared between patients but cannot be compared to other cell types in the same patient (i.e. inter-sample comparisons of the same cell type but not intra-individual comparisons of
different cell types). On the other hand, some cell population data can be compared to other cell populations within the analyzed patient but not the same cell population in a different
patient (i.e. intra-sample comparison between cell types). To validate these transcriptome-based data (Supplementary Table S2), they were compared to IHC or FCM data. Formalin-fixed
paraffin-embedded (FFPE) slides were stained with the indicated antibodies (Supplementary Fig. S1). Images were digitally captured and analyzed with Tissue Studio 2.0 to establish immune
cell numbers and calculate the immune cell densities (/mm2) (Supplementary Table S3). Tumor-infiltrating immune cells were also evaluated by FCM (Supplementary Table S4). Figure 1 summarizes
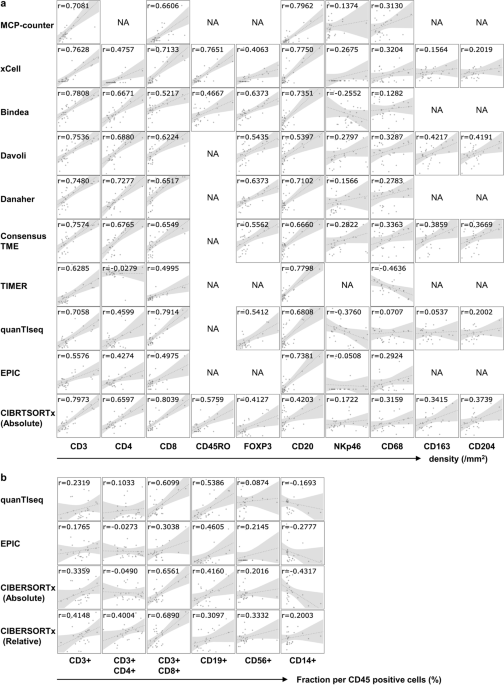
the results of bivariate Pearson correlations between the indicated computational transcriptome-based quantification methods and IHC/FCM data for the corresponding cells. The estimated
scores of T cells and B cells by the RNA-Seq-based methods correlated closely with the density of CD3-, CD4-, CD8-, and CD20-positive cells by IHC (Fig. 1a) and the fraction of CD3-, CD8-,
CD19- positive cells by FCM (Fig. 1b). Among them, the estimation of CD8-positive T cells in CIBERSORTx absolute mode exhibited the highest coefficient (0.8039). In addition, the
single-sample Gene Set Enrichment Analysis (ssGSEA) score using the gene set of Bindea et al. and Danaher et al. for FOXP3-positive regulatory T cells (0.6373) and the MCP-counter for
CD20-positive B cells (0.7962) also showed a high correlation coefficient with IHC. On the other hand, transcriptome-based quantification of NK cells (NKp46) or macrophages (CD68) displayed
less correlation with IHC; the highest correlation coefficient with NK cells (NKp46) and macrophages (CD68) was obtained by quanTIseq (0.3760) and TIMER (0.4636), respectively. For
transcriptome-based estimation, PBMC benchmarks may be less informative12. Therefore, we performed ssGSEA analysis using the gene set for tumor-associated macrophages (TAM) by Cassetta et
al.13. However, it could not improve the correlation with CD68, CD163 or CD204-positive cells (0.4148, 0.4708 or 0.4119, respectively, Supplementary Fig. S2). In Supplementary Fig. S3, the
transcriptome-based methods were ranked according to the correlation coefficient with the densities of immune cells. None of the methods outperformed others in every immune cell type.
SPATIAL AND FUNCTIONAL ANALYSIS OF TUMOR-INFILTRATING T CELLS Compared to IHC, spatial analysis by RNA-Seq is challenging. First, immune cell densities at the core of the tumor (CT) and
invasive margin (IM) of the tumor were enumerated in each patient (Supplementary Fig. S4 and Supplementary Table S5). Next, RNA-Seq data were subjected to Tumor Immune Dysfunction and
Exclusion (TIDE) framework approaches to compute a T cell exclusion score (Supplementary Table S6)14. Finally, the exclusion scores of 30 gastric cancer patients were compared to the ratio
of cell densities at the CT versus IM of the CD3+, CD4+ or CD8+ cells (Fig. 2a). TIDE exclusion scores negatively correlated with the ratio of CT/IM densities for CD8+ cells, suggesting that
RNA-Seq can be utilized to evaluate the spatial distribution of immune cells, particularly CD8+ T cells. Tumor-infiltrating cells (TICs) were isolated from the surgically resected tumors
and their capacity to produce IFN-γ, TNF-α and IL-2 was examined by FCM (Supplementary Fig. 5a and Table S7). TICs were left unstimulated or were stimulated with CytoStim (CS), which
stimulates T cells via the T cell receptor (TCR), or were stimulated with Phorbol 12-Myristate 13-Acetate/Ionomycin (PMA/IM; PI), which directly increases intracellular calcium concentration
without TCR signaling and results in cytokine expression (Supplementary Fig. S5b). Were TCR signaling to be suppressed by the immune inhibitory molecules, cytokine production following
CytoStim stimulation would be decreased relative to PMA/IM stimulation. Therefore, using FCM, the level of T cell dysfunction can be evaluated by the differences in the percentage of
cytokine-producing cells between PMA/IM stimulation versus CytoStim stimulation (PI-CS). As shown in Fig. 2b, TIDE dysfunction scores of the 30 patients correlated well with differences in
the percentages of cytokine-producing cells of PI-CS (Fig. 2b). PROLIFERATION OF TUMOR CELLS AND IMMUNE CELLS Ki-67 staining is a well-established method for detecting proliferating cells.
In the case of IHC, Ki-67+ tumor cells and immune cells can be discriminated morphologically (Fig. 3a, Supplementary Table S8). However, RNA-based evaluation methods for cell proliferation
cannot predict the type of proliferating cells. As shown in Fig. 3b, the densities of Ki-67+ tumor cells, Ki-67+ immune cells and all Ki-67+ cells (both tumor and immune cells) were closely
correlated with one another (Fig. 3b). These results suggest that extensively proliferating tumor cells are associated with proliferating immune cells. Therefore, we screened the appropriate
gene sets to reflect the proliferation of the tumor cells and the immune cells all together (Supplementary Table S9). The ssGSEA scores of DNA REPLICATION from the REACTOME subset of
canonical pathways in MSigDB (http://www.gsea-msigdb.org/gsea/msigdb/collections.jsp) displayed the highest correlation with the Ki-67-positive cell density by IHC (Fig. 3c). CYTOTOXIC
ACTIVITY IN THE TUMOR As shown in Fig. 3d, damaged tumor cells were detected and enumerated on histology slides (Supplementary Table S10). The correlation between the total cell death count
by histology and ssGSEA scores was examined to identify the appropriate gene sets for evaluating cell death in the tumor (Supplementary Table S11). GOBP_NECROPTOTIC_SIGNALING_PATHWAY from
MSigDB exhibited the highest correlation coefficient of 0.7231 (Fig. 3e). TME ANALYSIS WITH THE RELIABLE GENE SETS To obtain cell fractions that can be compared within samples (intra-sample
comparison), quanTIseq and CIBERSORTx are recommended. For inter-sample comparison of any cells of interest, we selected 7 reliable transcriptome-based parameters supported by histology and
FCM to evaluate the gastric cancer TME (Table 3 and Supplementary Fig. S3). In terms of the numbers and percentages of immune cells, the CIBERSORTx absolute mode for CD8+ T cells,
Bindea's and Danaher’s gene set analysis for regulatory T cells, and the MCP-counter for B cells were selected. IHC and FCM confirmed the appropriateness of using the TIDE framework for
evaluating the exclusion and dysfunction of immune cells in gastric cancer. The ssGSEA scores of REACTOME_DNA_REPLICATION and GOBP_NECROPTOTIC_SIGNALING_PATHWAY can be utilized for
evaluating proliferation and cell death in the tumor. With these 7 selected transcriptome-based methods, the inter-sample comparison of TME in 30 gastric cancer patients was performed. The
30 gastric cancer patients were first clustered into two groups; the Immune-Rich (IR) and the Immune-Poor (IP) groups (Fig. 4a). Scores for CD8+ T cells, Tregs and B cells were high in the
IR group, whereas the exclusion scores were consistently low. T cells became dysfunctional and both proliferation and cell death were evident in the IR patients. The IP group was further
divided into IP dysfunctional (IPd) and proliferative (IPp). Scores for Tregs, exclusion, and dysfunction were high in IPd, while scores for proliferation were high and dysfunction were low
in IPp. Patients with MSI and EBV subtypes (TCGA molecular classification) or “Hot” tumors (by Sato's immunogram classification8) were enriched in the IR group, while GS and CIN types
were enriched in the IP group. In addition, PD-L1 expression by tumor or immune cells was present in IR patients. In survival analysis, IPd patients had the worst overall survival (OS) (Fig.
4c, _P_ = 0.007, log-rank test). Three hundred seventy-five gastric cancer patients from the TCGA cohort were similarly clustered into IR, IPd and IPp groups (Fig. 4b and Supplementary
Table S12). Although the differences were not statistically significant, the worst OS in the IPd group was also detected (Fig. 4d, _P_ = 0.06, log-rank test). We compared the survival
analysis of the TCGA cohort between Sato’s immunogram classification8 and the transcriptome-based TME classification of this study. Eighty Hot1 cases were re-classified as 61 IR (Hot1-IR),
10 IPd (Hot1-IPd) and 9 IPp (Hot1-IPp) subtypes (Supplementary Fig. S6a). OS of Hot1-IR was better than that of Hot1, and OS of Hot1-IPd and Hot1-IPp was worse than that of Hot1
(Supplementary Fig. S6b). Similarly, 187 Cold cases were classified as 34 IR (Cold-IR), 38 IPd (Cold-IPd) and 115 IPp (Cold-IPp). OS of Cold-IR was better than that of Cold, while OS of
Cold-IPd and Cold-IPp was comparable to that of Cold patients (Supplementary Fig. S6e). DISCUSSION RNA-Seq data of bulk tumor tissues treats heterogeneous cell populations as a whole; data
are averages of different cells with various gene expression levels. Therefore, computational methods to evaluate cellular composition are essential. There are now many tools available for
this purpose16. In an earlier study, we selected gene sets to quantify immunological parameters in the TME by comparing them with similar gene sets available in the literature17. However,
there were some discrepancies between the quantification results of cell populations by transcriptomic analyses and the density of the corresponding cell type in a tissue. Therefore, in this
study, we applied several computational transcriptome analysis methods for evaluating the immune cell composition and immune-related TME of gastric cancer and compared the results to IHC
and FCM data, which are regarded as gold standards for TME analysis. Transcriptome-based quantification of CD8+ T cells, regulatory T cells and B cells in the tumor was quite reliable (Fig.
1). Furthermore, we demonstrated that even spatial and functional analysis is feasible by RNA-Seq (Fig. 2); the “Exclusion score” was validated by the absence of CD8+ T cells in the core of
the tumor as shown by IHC. The “Dysfunction score” was validated by the detection of fewer IFN-γ producing capacity of CD8+ T cells in the tumor using FCM. We also identified appropriate
gene sets that reflected cell proliferation or cell death in the tumor (Fig. 3). The output scores of the different methods allow either inter-sample comparisons of the same cell type,
intra-sample comparisons between different cell types, or both (Table 2). In fact, IHC using a single antibody allows inter-sample comparisons, while FCM is good for intra-sample
comparisons, depending on the panel of antibodies used. Therefore, transcriptome-based cell type quantification methods for inter-sample comparison were validated with IHC, and methods for
intra-sample comparison were compared to FCM (Fig. 1). QuanTIseq, EPIC, and the absolute mode of CIBERSORTx generate an absolute score that can be interpreted as a cell fraction. Therefore,
they can be utilized for both inter- and intra-sample comparisons and are quite useful in this context. Cell-type-specific estimation in the TME using bulk tumor data is challenging.
Predicting transcriptionally distinct cell types would likely show good correlations. For example, Bindea et al. and Danaher et al. took FOXP3 as a single marker gene for Treg and
successfully hit the best correlation coefficient of 0.6373 (Fig. 1). However, gene expression levels change according to their activation and differentiation status in many other cell
types. Therefore, the gene set approach was used to address these complex problems as the wisdom of the crowd18. Although different gene sets were proposed by different methods, strategies
to assemble gene sets for immune cells, in general, depend on the expression profiles of purified cell types to identify reference genes and therefore rely heavily on the data source from
which the references are inferred and could this be inclined to overfitting these data. Therefore, it is difficult to cover the estimation of all cell types with a single prediction tool
(Supplementary Fig. S3). The difficulties in quantifying NK cells and macrophages are not limited to transcriptome-based methods. Because NK cells and macrophages express different arrays of
cell surface receptors, the expression of which overlap in different cell populations, identification and quantification of these cells is not possible by a single marker assay19,20. For
example, macrophage marker CD68 is also expressed by γδ T cells, NK cells, a subset of B cells, fibroblasts, and endothelial cells. CD163 is also expressed by dendritic cells. CD204 is
expressed by only a subset of M2 macrophages. NKp46 is expressed by CD56bright NK cells, but not CD56dim NK cells19. Ki67 staining is used as a standard for evaluating proliferating cells.
However, Ki67 protein levels were not a simple on-and-off switch of cell proliferation. Because Ki67 protein is continuously produced from the start of S phase and Ki67 is continuously
degraded during G0 and G1 phase, quiescent cells re-entering the cell cycle will have varying levels of Ki6721. The discrepancy between transcriptome-based methods and IHC for NK cells,
macrophages and cell proliferation might be due to the uncertainty of IHC results rather than the ambiguity of transcriptome-based methods. Multiplex IHC will overcome this problem and can
be used as the gold standard for such assays. IHC and FCM have been used as gold standards to estimate the immune cell content within a sample10. However, there are several limitations to
these methods. Only a limited number of cell-type-specific markers can be utilized. FCM requires a large amount of sample that should be mechanically or enzymatically dissociated to isolate
single-cell suspensions. Different single-cell dissociation efficiencies might bias the apparent proportions of immune cells in the tumor. In contrast, gene expression profiling by RNA-Seq
provides comprehensive transcriptomics datasets derived from small tumor samples, and a large number of markers can be analyzed simultaneously. In addition to inflammatory molecules, many
biological processes that shape the TME, such as angiogenesis, metabolism, and response to hypoxia, can be assessed and incorporated into the marker panel. Although we incorporated only 7
parameters that were validated by IHC or FCM into the TME analysis in this study, transcriptome-based TME analysis can easily be integrated with other molecular analyses and extended in the
future. Using 7 parameters extracted from RNA-Seq of bulk tissues, gastric cancers were immunologically classified into 3 clusters (Fig. 4). As reported previously, patients whose tumors had
an immunologically “hot” TME had a better post-surgical prognosis8. Similar results were obtained in the current study (Fig. 4), although different gene sets and algorithms were applied. In
Fig. 4a, two MSI cases, BKT008 and 038, were classified as belonging to the IP group, while the other 3 MSI cases were clustered in the IR group. The prognosis of these two IP patients was
poor; BKT008 died of the primary disease 741 days after surgery and BKT038 after 347 days (Supplementary Tables S1 and S13). A poor immune response in the tumor might be responsible for the
shorter OS of these two patients, despite their MSI subtype. In addition, BKT053, which was classified as CIN by TCGA classification and Hot1 by immunogram classification, was re-classified
as IPp in this study. BKT053 relapsed on day 223 and died on 482 days. As shown in Supplementary Fig. S6a, transcriptome-based TME classification can discriminate IP patients from Hot1
patients in the TCGA cohort. Immunogram classification was based on the concept of the cancer-immunity cycle7, and TME classification was more associated with intratumoral immune response.
Both methods are not mutually exclusive. Combining these two classifications can fine-tune the immunological subtypes of gastric cancer. In conclusion, computational methods for
transcriptomic analysis were validated by comparison with IHC and FCM to evaluate the TME of gastric cancer. It is feasible to evaluate the TME using RNA-Seq data obtained from small bulk
tissues. For intra-sample comparison, either quanTIseq or CIBERSORTx is an appropriate tool to evaluate the immune cell fractions in TME. For inter-sample comparison, selecting the best
score method for each parameter is better than any single method. Using 7 parameters selected, the TME of gastric cancer could be appropriately immunologically classified. METHODS PATIENTS
AND DATA SETS We enrolled 30 patients who underwent gastrectomy at Tokyo Metropolitan Bokutoh Hospital between June 2014 and October 2017 (Table 1). Clinical profiles with histology by the
Lauren classification, overexpression of human epidermal growth factor receptor 2 (HER2) protein and the presence or absence of _Helicobacter pylori_ infection were reported in our previous
work8. In the previous study, RNA-Seq of bulk tumor tissue was performed8. RNA-Seq data of BKT patients are available at DDBJ Sequence Read Archive (Accession no. DRA009379)8.
Clinicopathological features and RNA-Seq data for 375 additional gastric cancer patients were downloaded from the TCGA portal site (https://portal.gdc.cancer.gov/). COMPUTATIONAL METHODS TO
ANALYZE RNA-SEQ DATA RNA-Seq data were analyzed using the following algorithms or web tools: MCP-counter ("MCPcounter" R package)22, xCell (https://xcell.ucsf.edu/)23, TIMER
(http://timer.cistrome.org/)24, quanTlseq (https://icbi.i-med.ac.at/software/quantiseq/doc/)25, EPIC (http://epic.gfellerlab.org)26, and CIBERSORTx (https://cibersortx.stanford.edu/)27.
ssGSEA was performed using the gene sets of Bindea28, Davoli29, Danaher30, Cassetta13 and Consensus TME31 to enumerate immune cell composition. The enrichment scores were obtained using the
ssGSEA method with R package ssGSEA 2.0 (https://github.com/broadinstitute/ssGSEA2.0) and R software version 3.6.0. The list of genes used by each method is provided in Supplementary Table
S14. Because of the variation in the degree of specificity to which cell subsets were defined, summations of subsets were required to allow accurate comparisons in each cell type
(Supplementary Tables S15 and S16). T cell dysfunction and exclusion scores were calculated on the TIDE website (http://tide.dfci.harvard.edu/)14. TCGA molecular classification of gastric
cancer was performed as previously reported6,8. Gastric cancers were classified as EBV, MSI, genomic stability (GS), or chromosomal instability (CIN). Gastric cancers were also grouped as
Mesenchymal or Non‐Mesenchymal subtypes by their 71‐gene mesenchymal signature according to the ACRG project32. Immunological subtypes based on immunogram scores were determined in our
previous work8. HISTOLOGICAL ANALYSIS FFPE blocks were obtained from the Pathology Department of Tokyo Metropolitan Bokutoh Hospital. Immunohistochemistry was performed using the Ventana
BenchMark automated immunostainer (Ventana Medical Systems, Tucson, AZ, USA) with labeled streptavidin–biotin and visualized with 3,3′-diaminobenzidine. The primary antibodies used were
anti-CD3 (clone LN10, Leica), -CD4 (clone SP35, Ventana), -CD8 (clone 4B11, Leica), -CD45RO (clone UCHL-1, Ventana), -FOXP3 (clone 236A/E7, Abcam), -CD20 (clone L26, Leica), -NKp46 (clone
#195314, R&D), -CD68 (clone Kp-1, Dako), -CD163 (clone 10D6, Leica), -CD204 (clone SRA-E5, Transgenic), -Ki-67 (clone MIB-1, Dako), -PD-L1 (clone E1L3N, Cell Signaling), -MLH1 (clone
ES05, Leica), -MSH2 (clone FE11, Dako), -MSH6 (clone Polyclonal (Rabbit), GeneTex) and -PMS2 (clone M0R4G, Leica). EBV-encoded small RNA in situ hybridization (EBER-ISH) was performed on
paraffin sections using a fluorescein isothiocyanate (FITC)-labeled peptide nucleic acid probe (Y5200; Dako, Glostrup, Denmark) and anti-FITC antibody (V0403, Dako). Slides were digitized
with a Nanozoomer 2.0-HT virtual slide scanner (Hamamatsu Photonics, Hamamatsu, Japan) and observed in the NDP.view2 software (Hamamatsu Photonics). The density of immune cells was analyzed
by Tissue Studio 2.0 software (Definiens, Munich, Germany). FLOW CYTOMETRY Tumors were cut into small pieces and enzymatically dissociated using a tumor dissociation kit (Miltenyi Biotec
Inc., Auburn, CA, USA) to prepare fresh tumor digest (FTD) according to the manufacturer's instructions. After passing through a 70‐μm cell strainer (Thermo Fisher Scientific, Hampton,
NH, USA), FTDs were cryopreserved in Bambanker™ freezing medium (NIPPON Genetics, Tokyo, Japan) until analysis. Cryopreserved FTDs were thawed in RPMI, and then stained using a Zombie Aqua™
Fixable Viability Kit (BioLegend, San Diego, CA, USA) with anti‐CD45 (clone 2D1, BioLegend) and -CD3 (clone HIT3a, BioLegend), -CD4 (clone SK3, Thermo Fisher Scientific), -CD8 (clone, HIT8a,
BioLegend), -CD14 (clone: M5E2, BioLegend), -CD19 (clone J3-119, Beckman Coulter), -CD56-PE (clone N901, Beckman Coulter), -IFN‐γ (clone 45.15, Beckman Coulter, Brea, CA, USA), -TNF‐α
(clone MAb11, BioLegend), -IL‐2 (clone MQ1-17H12, BioLegend). For the detection of cytokine production, cells were stimulated with 10 ng/ml Phorbol 12‐Myristate 13‐Acetate (PMA;
Sigma‐Aldrich, St. Louis, MO, USA) together with 1 μg/ml Ionomycin (IM; Sigma‐Aldrich) or CytoStim (CS; Miltenyi Biotec) in the presence of 10 µg/ml brefeldin A (Sigma‐Aldrich) at 37ºC for 4
h. Intracellular cytokine staining was then carried out according to the manufacturer's instructions (using IntraPrep Permeabilization Reagent; Beckman Coulter). Stained cells were
analyzed on a Gallios flow cytometer (Beckman Coulter) and data were processed using Kaluza (Beckman Coulter) and FlowJo (version 7.6.5; TreeStar, Ashland, OR, USA) software. STATISTICAL
ANALYSES For analyzing the correlation between each factor, Pearson's correlation coefficient method was used. For categorical variables, the chi-square test was used. A heat map was
created using Ward’s hierarchical cluster analysis. The Kaplan–Meier method was used for survival analysis, and comparisons between groups were performed by log-rank testing. JMP Pro 15 (SAS
Institute Japan, Tokyo, Japan) was used for statistical analysis. A value of _P_ < 0.05 was considered statistically significant. ETHICAL DECLARATIONS This study was approved by the
Research Ethics Committees of the University of Tokyo (No. G3545) and Tokyo Metropolitan Bokutoh Hospital (No. 25‐38‐02). All procedures followed were in accordance with the ethical
standards of the responsible committee on human experimentation (institutional and national) and with the Helsinki Declaration of 1964 and later versions. Informed written consent was
obtained from all patients included in the study. DATA AVAILABILITY Data are deposited on DDBJ Sequence Read Archive (Accession no. DRA009379). REFERENCES * Sung, H. _et al._ Global cancer
statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. _CA Cancer J. Clin._ 71, 209–249. https://doi.org/10.3322/caac.21660 (2021). Article
PubMed Google Scholar * Kang, Y. K. _et al._ Nivolumab in patients with advanced gastric or gastro-oesophageal junction cancer refractory to, or intolerant of, at least two previous
chemotherapy regimens (ONO-4538–12, ATTRACTION-2): A randomised, double-blind, placebo-controlled, phase 3 trial. _Lancet_ 390, 2461–2471. https://doi.org/10.1016/s0140-6736(17)31827-5
(2017). Article CAS PubMed Google Scholar * Fuchs, C. S. _et al._ Safety and efficacy of pembrolizumab monotherapy in patients with previously treated advanced gastric and
gastroesophageal junction cancer: Phase 2 clinical KEYNOTE-059 trial. _JAMA Oncol._ 4, e180013. https://doi.org/10.1001/jamaoncol.2018.0013 (2018). Article PubMed PubMed Central Google
Scholar * Shitara, K. _et al._ Pembrolizumab versus paclitaxel for previously treated, advanced gastric or gastro-oesophageal junction cancer (KEYNOTE-061): A randomised, open-label,
controlled, phase 3 trial. _Lancet_ 392, 123–133. https://doi.org/10.1016/s0140-6736(18)31257-1 (2018). Article CAS PubMed Google Scholar * Shitara, K. _et al._ Efficacy and safety of
pembrolizumab or pembrolizumab plus chemotherapy vs chemotherapy alone for patients with first-line, advanced gastric cancer: The KEYNOTE-062 phase 3 randomized clinical trial. _JAMA Oncol._
6, 1571–1580. https://doi.org/10.1001/jamaoncol.2020.3370 (2020). Article PubMed Google Scholar * TCGA. Comprehensive molecular characterization of gastric adenocarcinoma. _Nature_ 513,
202–209. https://doi.org/10.1038/nature13480 (2014). Article ADS CAS Google Scholar * Chen, D. S. & Mellman, I. Oncology meets immunology: The cancer-immunity cycle. _Immunity_ 39,
1–10. https://doi.org/10.1016/j.immuni.2013.07.012 (2013). Article CAS PubMed Google Scholar * Sato, Y. _et al._ Integrative immunogenomic analysis of gastric cancer dictates novel
immunological classification and the functional status of tumor-infiltrating cells. _Clin. Transl. Immunol._ 9, e1194. https://doi.org/10.1002/cti2.1194 (2020). Article CAS Google Scholar
* Kim, S. T. _et al._ Comprehensive molecular characterization of clinical responses to PD-1 inhibition in metastatic gastric cancer. _Nat. Med._ 24, 1449–1458.
https://doi.org/10.1038/s41591-018-0101-z (2018). Article CAS PubMed Google Scholar * Petitprez, F. _et al._ Quantitative analyses of the tumor microenvironment composition and
orientation in the era of precision medicine. _Front. Oncol._ 8, 390. https://doi.org/10.3389/fonc.2018.00390 (2018). Article PubMed PubMed Central Google Scholar * Bang, Y. J. _et al._
Trastuzumab in combination with chemotherapy versus chemotherapy alone for treatment of HER2-positive advanced gastric or gastro-oesophageal junction cancer (ToGA): A phase 3, open-label,
randomised controlled trial. _Lancet_ 376, 687–697. https://doi.org/10.1016/s0140-6736(10)61121-x (2010). Article CAS PubMed Google Scholar * Baine, M. J. _et al._ Transcriptional
profiling of peripheral blood mononuclear cells in pancreatic cancer patients identifies novel genes with potential diagnostic utility. _PLoS ONE_ 6, e17014.
https://doi.org/10.1371/journal.pone.0017014 (2011). Article ADS CAS PubMed PubMed Central Google Scholar * Cassetta, L. _et al._ Human tumor-associated macrophage and monocyte
transcriptional landscapes reveal cancer-specific reprogramming, biomarkers, and therapeutic targets. _Cancer Cell_ 35, 588.e510-602.e510. https://doi.org/10.1016/j.ccell.2019.02.009 (2019).
Article CAS Google Scholar * Jiang, P. _et al._ Signatures of T cell dysfunction and exclusion predict cancer immunotherapy response. _Nat. Med._ 24, 1550–1558.
https://doi.org/10.1038/s41591-018-0136-1 (2018). Article CAS PubMed PubMed Central Google Scholar * Oh, S. C. _et al._ Clinical and genomic landscape of gastric cancer with a
mesenchymal phenotype. _Nat. Commun._ 9, 1777. https://doi.org/10.1038/s41467-018-04179-8 (2018). Article ADS CAS PubMed PubMed Central Google Scholar * Finotello, F., Rieder, D.,
Hackl, H. & Trajanoski, Z. Next-generation computational tools for interrogating cancer immunity. _Nat. Rev. Genet._ 20, 724–746. https://doi.org/10.1038/s41576-019-0166-7 (2019).
Article CAS PubMed Google Scholar * Kobayashi, Y., Kushihara, Y., Saito, N., Yamaguchi, S. & Kakimi, K. A novel scoring method based on RNA-Seq immunograms describing individual
cancer-immunity interactions. _Cancer Sci._ 111, 4031–4040. https://doi.org/10.1111/cas.14621 (2020). Article CAS PubMed PubMed Central Google Scholar * Marbach, D. _et al._ Wisdom of
crowds for robust gene network inference. _Nat. Methods_ 9, 796–804. https://doi.org/10.1038/nmeth.2016 (2012). Article CAS PubMed PubMed Central Google Scholar * Freud, A. G.,
Mundy-Bosse, B. L., Yu, J. & Caligiuri, M. A. The broad spectrum of human natural killer cell diversity. _Immunity_ 47, 820–833. https://doi.org/10.1016/j.immuni.2017.10.008 (2017).
Article CAS PubMed PubMed Central Google Scholar * Wu, K. _et al._ Redefining tumor-associated macrophage subpopulations and functions in the tumor microenvironment. _Front. Immunol._
https://doi.org/10.3389/fimmu.2020.01731 (2020). Article PubMed PubMed Central Google Scholar * Miller, I. _et al._ Ki67 is a graded rather than a binary marker of proliferation versus
quiescence. _Cell Rep._ 24, 1105.e1105-1112.e1105. https://doi.org/10.1016/j.celrep.2018.06.110 (2018). Article CAS Google Scholar * Becht, E. _et al._ Estimating the population abundance
of tissue-infiltrating immune and stromal cell populations using gene expression. _Genome Biol._ 17, 218. https://doi.org/10.1186/s13059-016-1070-5 (2016). Article CAS PubMed PubMed
Central Google Scholar * Aran, D., Hu, Z. & Butte, A. J. xCell: Digitally portraying the tissue cellular heterogeneity landscape. _Genome Biol._ 18, 220.
https://doi.org/10.1186/s13059-017-1349-1 (2017). Article CAS PubMed PubMed Central Google Scholar * Li, T. _et al._ TIMER: A web server for comprehensive analysis of tumor-infiltrating
immune cells. _Cancer Res._ 77, e108–e110. https://doi.org/10.1158/0008-5472.Can-17-0307 (2017). Article CAS PubMed PubMed Central Google Scholar * Finotello, F. _et al._ Molecular and
pharmacological modulators of the tumor immune contexture revealed by deconvolution of RNA-seq data. _Genome Med._ 11, 34. https://doi.org/10.1186/s13073-019-0638-6 (2019). Article CAS
PubMed PubMed Central Google Scholar * Racle, J., de Jonge, K., Baumgaertner, P., Speiser, D. E. & Gfeller, D. Simultaneous enumeration of cancer and immune cell types from bulk tumor
gene expression data. _Elife_ https://doi.org/10.7554/eLife.26476 (2017). Article PubMed PubMed Central Google Scholar * Newman, A. M. _et al._ Determining cell type abundance and
expression from bulk tissues with digital cytometry. _Nat. Biotechnol._ 37, 773–782. https://doi.org/10.1038/s41587-019-0114-2 (2019). Article CAS PubMed PubMed Central Google Scholar *
Bindea, G. _et al._ Spatiotemporal dynamics of intratumoral immune cells reveal the immune landscape in human cancer. _Immunity_ 39, 782–795. https://doi.org/10.1016/j.immuni.2013.10.003
(2013). Article CAS PubMed Google Scholar * Davoli, T., Uno, H., Wooten, E. C. & Elledge, S. J. Tumor aneuploidy correlates with markers of immune evasion and with reduced response
to immunotherapy. _Science_ https://doi.org/10.1126/science.aaf8399 (2017). Article PubMed PubMed Central Google Scholar * Danaher, P. _et al._ Gene expression markers of tumor
infiltrating leukocytes. _J. Immunother. Cancer_ 5, 18. https://doi.org/10.1186/s40425-017-0215-8 (2017). Article PubMed PubMed Central Google Scholar * Jiménez-Sánchez, A., Cast, O.
& Miller, M. L. Comprehensive benchmarking and integration of tumor microenvironment cell estimation methods. _Cancer Res._ 79, 6238–6246. https://doi.org/10.1158/0008-5472.Can-18-3560
(2019). Article PubMed Google Scholar * Lee, J. _et al._ Development of mesenchymal subtype gene signature for clinical application in gastric cancer. _Oncotarget_ 8, 66305–66315.
https://doi.org/10.18632/oncotarget.19985 (2017). Article PubMed PubMed Central Google Scholar Download references ACKNOWLEDGEMENTS The authors thank Mikiko Shibuya, Yaeko Furuhashi,
Yuki Yoshikawa for excellent technical assistance. FUNDING This work was supported in part by AMED under Grant Number JP 19cm0106552 and JP 21ck0106639. AUTHOR INFORMATION AUTHORS AND
AFFILIATIONS * Department of Gastrointestinal Surgery, The University of Tokyo Graduate School of Medicine, Tokyo, 113-8655, Japan Noriyuki Saito & Yasuyuki Seto * Department of
Immunotherapeutics, The University of Tokyo Hospital, 7-3-1 Hongo, Bunkyo-Ku, Tokyo, 113-8655, Japan Noriyuki Saito, Yukari Kobayashi, Koji Nagaoka, Yoshihiro Kushihara & Kazuhiro Kakimi
* Department of Medical Oncology, The Cancer Institute Hospital of Japanese Foundation for Cancer Research, Tokyo, 135-8550, Japan Yasuyoshi Sato * Department of Pathology, Graduate School
of Medicine, The University of Tokyo, Tokyo, Japan Hiroyuki Abe & Tetsuo Ushiku * Department of Surgery, Tokyo Metropolitan Bokutoh Hospital, Tokyo, 130-8575, Japan Ikuo Wada Authors *
Noriyuki Saito View author publications You can also search for this author inPubMed Google Scholar * Yasuyoshi Sato View author publications You can also search for this author inPubMed
Google Scholar * Hiroyuki Abe View author publications You can also search for this author inPubMed Google Scholar * Ikuo Wada View author publications You can also search for this author
inPubMed Google Scholar * Yukari Kobayashi View author publications You can also search for this author inPubMed Google Scholar * Koji Nagaoka View author publications You can also search
for this author inPubMed Google Scholar * Yoshihiro Kushihara View author publications You can also search for this author inPubMed Google Scholar * Tetsuo Ushiku View author publications
You can also search for this author inPubMed Google Scholar * Yasuyuki Seto View author publications You can also search for this author inPubMed Google Scholar * Kazuhiro Kakimi View author
publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS N.S. conducted the experiments, analyzed and interpreted the data, prepared the figures and wrote the
manuscript. Y.Sa. designed the study, analyzed and interpreted the data, H.A. and T.U. provided methodological expertise and performed immunohistochemistry. I.W. collected samples and
analyzed the data. Y.Ko. provided methodological expertise and performed flowcytometry. K.N. and Y.Ku. provided methodological expertise and performed RNA-Seq. Y.Se. supervised the study,
and interpreted the results. K.K. designed the study, analyzed and interpreted the data, and wrote the manuscript. All authors reviewed the manuscript. CORRESPONDING AUTHOR Correspondence to
Kazuhiro Kakimi. ETHICS DECLARATIONS COMPETING INTERESTS Dr. Kakimi reports Grants from TAKARA BIO Inc. outside the submitted work. The Department of Immunotherapeutics, The University of
Tokyo Hospital, is endowed by TAKARA BIO Inc. The other authors have no conflict of interests to disclose. ADDITIONAL INFORMATION PUBLISHER'S NOTE Springer Nature remains neutral with
regard to jurisdictional claims in published maps and institutional affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY FIGURES. SUPPLEMENTARY TABLES. RIGHTS AND PERMISSIONS OPEN ACCESS
This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as
long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third
party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the
article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the
copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Saito, N., Sato, Y., Abe,
H. _et al._ Selection of RNA-based evaluation methods for tumor microenvironment by comparing with histochemical and flow cytometric analyses in gastric cancer. _Sci Rep_ 12, 8576 (2022).
https://doi.org/10.1038/s41598-022-12610-w Download citation * Received: 28 December 2021 * Accepted: 10 May 2022 * Published: 20 May 2022 * DOI: https://doi.org/10.1038/s41598-022-12610-w
SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to
clipboard Provided by the Springer Nature SharedIt content-sharing initiative