Yy1’s role in the peg3 imprinted domain
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT The ICR (Imprinting Control Region) of the _Peg3_ (Paternally Expressed Gene 3) domain contains an unusual cluster of YY1 binding sites. In the current study, these YY1 binding
sites were mutated to characterize the unknown roles in the mouse _Peg3_ domain. According to the results, paternal and maternal transmission of the mutant allele did not cause any major
effect on the survival of the pups. In the mutants, the maternal-specific DNA methylation on the ICR was properly established and maintained, causing no major effect on the imprinting of the
domain. In contrast, the paternal transmission resulted in changes in the expression levels of several genes: down-regulation of _Peg3_ and _Usp29_ and up-regulation of _Zim1_. These
changes were more pronounced during the neonatal stage than during the adult stage. In the case of _Peg3_ and _Zim1_, the levels of the observed changes were also different between males and
females, suggesting the different degrees of YY1 involvement between two sexes. Overall, the results indicated that YY1 is mainly involved in controlling the transcriptional levels, but not
the DNA methylation, of the _Peg3_ domain. SIMILAR CONTENT BEING VIEWED BY OTHERS ORIENTATION OF MOUSE _H19_ ICR AFFECTS IMPRINTED _H19_ GENE EXPRESSION THROUGH PROMOTER
METHYLATION-DEPENDENT AND -INDEPENDENT MECHANISMS Article Open access 17 December 2021 BALANCED GENE DOSAGE CONTROL RATHER THAN PARENTAL ORIGIN UNDERPINS GENOMIC IMPRINTING Article Open
access 29 July 2022 IDENTIFYING REGULATORS OF PARENTAL IMPRINTING BY CRISPR/CAS9 SCREENING IN HAPLOID HUMAN EMBRYONIC STEM CELLS Article Open access 18 November 2021 INTRODUCTION In mammals,
a small subset of genes are functionally different between two alleles due to genomic imprinting, by which one allele is preferentially expressed in a parental-origin-specific manner1.
These imprinted genes are usually clustered in specific regions of chromosomes, forming imprinted domains. Within a given domain, a small genomic region termed imprinting control region
(ICR) is responsible for inheriting germ cell-driven DNA methylation as a gametic signal, and also for controlling the transcription of the entire domain as a _cis_-regulatory region2. Two
DNA-binding proteins, CTCF (CCCTC-binding factor) and YY1 (Yin-yang 1), are known to bind to ICRs: CTCF to the ICR of the _H19_/_Igf2_ (Insulin-like growth factor 2) domain and YY1 to the
ICRs of the _Peg3_ and _Gnas_ (stimulatory G-protein alpha subunit) domains3,4,5,6. The DNA-binding sites of CTCF and YY1 within these ICRs are quite unusual. The number of these binding
sites found within an ICR ranges from 4 to 10, which is much greater than the number in the other regulatory regions, such as enhancers and promoters. These binding sites in a given ICR are
also all localized in a same orientation7. In the case of the ICR of _H19_/_Igf2_, the CTCF binding sites function as a methylation-sensitive insulator controlling the accessibility of the
promoters of both _H19_ and _Igf2_ to the shared enhancers located in the 3′-side of _H19_ 8. These CTCF binding sites are also required for maintaining the allele-specific DNA methylation
of the ICR. Either loss of the CTCF sites or reduced levels of the CTCF protein during early embryogenesis resulted in DNA methylation on the active allele of the ICR, demonstrating critical
roles played by CTCF for the functions of the ICR9,10,11,12. The YY1 binding sites localized within the ICR of the _Peg3_ domain have also been investigated through a series of _in vitro_
and _in vivo_ experiments. According to the results, reduced levels of the YY1 protein often resulted in changes in the DNA methylation levels of this ICR. Interestingly, the reduced levels
of YY1 usually caused DNA hypomethylation in the ICR during oogenesis and also in cell lines and somatic tissues13,14,15. On the other hand, the YY1 binding sites have been shown to function
as activators or repressors for the transcription of the _Peg3_ domain depending upon the testing systems and also the functional contexts. For instance, a series of reporter assays using
_in vitro_ systems revealed that the transcriptional activity of the bidirectional promoter of _Peg3_/_Usp29_ (Ubiquitin-specific protease 29) fluctuates, either up or down, depending upon
the number of the YY1 binding sites involved in the activity16. In contrast, the reduced levels of YY1 protein usually resulted in the increased transcriptional levels of _Peg3_, suggesting
that the YY1 binding sites might be repressors for the transcription of the _Peg3_ domain13,14,15, 17. However, the interpretation of these results has not been straightforward since
reducing the protein levels of YY1 might have impacted the _Peg3_ locus in both direct and indirect ways, especially through the other loci that are also controlled through YY1. To better
understand the _in vivo_ role of YY1 binding sites, thus, we have generated a mouse line carrying the mutated version of the 7 YY1 binding sites that are localized within the ICR of the
_Peg3_ domain. According to the results, these YY1 binding sites are not involved in the establishment and maintenance of the maternal-specific DNA methylation of the ICR. However, the YY1
binding sites appear to function as an activator for the transcription of the two adjacent genes, _Peg3_ and _Usp29_. Interestingly, the transcriptional level changes observed from this
mutant line were different between males and females, further suggesting the different degrees of YY1 involvement in the _Peg3_ domain between the two sexes. RESULTS GENERATION OF AN ALLELE
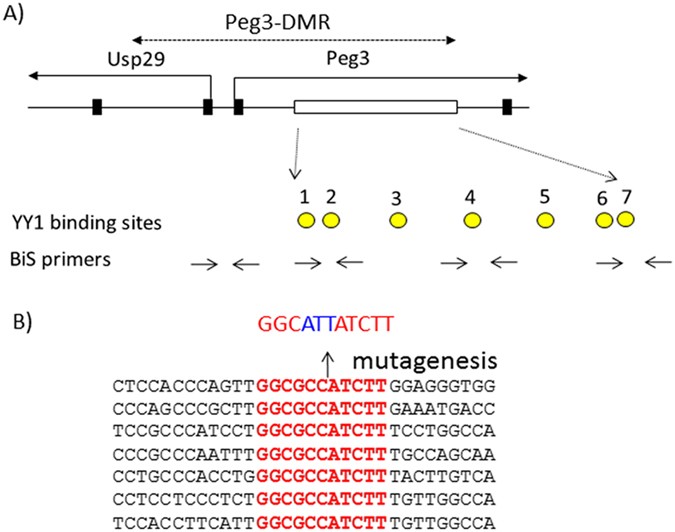
MUTATING THE 7 YY1 BINDING SITES WITHIN THE PEG3-DMR The ICR of the _Peg3_ domain corresponds to the 4-kb genomic region encompassing the 1.5-kb bidirectional promoter of _Peg3_/_Usp29_ and
the 2.5-kb 1st intron region of _Peg3_ (Fig. 1). This ICR has been also termed the Peg3-DMR (Differentially Methylated Region) given its allele-specific DNA methylation pattern5. The 7 YY1
binding sites within the 2.5-kb intron region were previously mutated for a series of _in vitro_ reporter assays16. The three bases of each binding site was changed from 5′-GCC-3′ to
5′-ATT-3′, which is part of the core binding motif for YY118. The 2.5-kb region containing the mutant version of 7 YY1 binding sites has been used to construct a targeting vector for mouse
knockout (KO) experiments (Fig. 2A). The final targeting vector was transfected into the ES cells of 129/SvJ. Transfected ES cells were subsequently screened with long-distance PCR and
southern blotting, identifying 20 targeted clones out of 300 ES cells (Fig. 2B,C). Two independent clones with the proper targeting were injected into the blastocysts of C57BL/6 J,
subsequently generating 10 chimeras. Among these chimeras, two were able to generate F1 pups with the germline transmission of the targeted allele. The F1 mice were further bred with a
Flippase line to remove the _NeoR_ (Neomycin Resistance) cassette, which is located between the 1.5-kb bidirectional promoter and the 2.5-kb intron regions. The 3-bp-mutation, 5′-GCC-3′ to
5′-ATT-3′, in each of the 7 YY1 binding sites was confirmed again through sequencing of the genomic DNA isolated from the F1 mice (data not shown). Besides the mutated 7 YY1 binding sites,
the final mutant allele also contains two loxP sites flanking the 2.5-kb intron region. Thus, a set of primers amplifying the 379-bp region surrounding the 2nd loxP site was used to genotype
all the progeny from the current study (Fig. 2D). Overall, we have successfully generated a mouse allele containing the mutant version of the 7 YY1 binding sites for the ICR of the _Peg3_
domain. PHENOTYPIC EFFECTS OF THE MUTANT ALLELE AT THE ORGANISM LEVEL The phenotypic effects of the mutant allele were analyzed at the organism level using a series of breeding experiments
(Supplemental material 1). Male and female heterozygotes were bred individually with their female and male wild-type littermates. The subsequent pups with the paternal and maternal
transmission of the mutant allele were analyzed in terms of their sex, genotype and body weight. The results are summarized as follows. First, the average litter size of the pups with the
paternal transmission of the mutant allele was smaller than the average litter size of the 129/B6 mice (7.76 vs 10), yet the ratio between the wild-type and heterozygote was close to the
mendelian ratio (WT:KO = 28:26), indicating no major embryonic lethality associated with the mutant allele. This is also the case for the maternal transmission of the mutant allele. The
average litter size was 7.6, and the ratio between the wild-type to heterozygote was also close to the mendelian ratio (WT:KO = 21:25). Second, the weight profiles of one-day-old neonates
also showed no major differences between WT and KO in both the paternal and maternal transmission (Supplemental material 1). Interestingly, the male heterozygotes from the paternal
transmission were slightly heavier than those from the wild-type littermates (104% with 12% S.D. versus 100% with 7% S.D., respectively), but this observation was not statistically
significant (Student t-test, _p_ = 0.075). It is also prudent to note that the mutant strain used for this study is the mixed background 129/B6. Thus, the outcome needs to be interpreted
with caution. Overall, the mutation on the 7 YY1 binding sites did not cause any major effects on the survival and growth rates of the mice. MUTATIONAL EFFECTS ON DNA METHYLATION LEVELS
Potential effects of the mutation on the DNA methylation levels of the Peg3-DMR were analyzed in the following manner. First, we isolated the DNA from the sperm that had been purified from
three sets of adult males. Each set has two heterozygotes and wild-type littermates. The first two sets were from 3-month-old mice, whereas the third set from 6-month-old mice. The isolated
DNA were first treated with the bisulfite conversion protocol, and later used as templates for PCR to amplify several target regions within the Peg3-DMR (Figs 1 and 3). We targeted three YY1
binding sites (Yy1-bs-2, -4, -7) as well as the bidirectional promoter region of _Peg3_/_Usp29_ (mPeg3-pro) (Fig. 3). We have also included the ICR of the _H19_/_Igf2_ domain (mH19) as an
internal control to monitor the purity of the isolated sperm (Fig. 3). The amplified PCR product from each locus was analyzed with COBRA (COmbined Bisulfite and Restriction Analysis19), and
later by individual sequencing (Fig. 3B). The sperm from the two sets of 3-month-old mice did not show any major difference between the wild-type and heterozygotes, showing no methylation in
the Peg3-DMR. However, the sperm from the 6-month-old heterozygote mice showed about 50% methylation at two regions, Yy1-bs-4 and -7, but not from the sperm of the wild-type littermates.
The results from individual sequencing also revealed that DNA methylation was detected exclusively from the mutant allele, but not from the wild-type allele, of the heterozygotes (Fig. 4B).
This suggests that the intact YY1 binding sites may be required for protecting DNA methylation during the spermatogenesis of the older mice. Second, we also repeated a similar series of DNA
methylation analyses using the mature eggs that had been isolated from three sets of females. Each set was comprised of three wild-type and heterozygote females of the 3-month-old age. In a
given set, we successfully obtained 60–80 eggs for each of the two groups, wild-type and heterozygote, which were subsequently used for DNA methylation analyses (Fig. 3). The results
revealed no major difference between the wild-type and heterozygote, both showing proper DNA methylation on the Peg3-DMR. Individual sequencing further indicated that the two alleles within
the heterozygotes also showed similar levels of DNA methylation between the wild-type and mutant alleles (Fig. 3B). Thus, this further confirmed that the YY1 binding sites may not be
required for the establishment of oocycte-specific DNA methylation on the Peg3-DMR. Third, we also performed a series of COBRA using the DNA isolated from the individual tissues of the adult
mice (Supplemental material 2). However, we did not observe any difference in the DNA methylation levels of the Peg3-DMR between the wild-type and heterozygote, confirming again that the
YY1 binding sites may be dispensable for the maintenance of the allele-specific DNA methylation of the Peg3-DMR. This has been further confirmed through a series of imprinting tests using
the hybrids that had been derived from the crossing of the male heterozygote of the 129/B6 background with the female breeders of the PWD/PhJ strain (Supplemental material 3). No change was
observed in the imprinting status of the genes in the _Peg3_ domain. Taken together, this series of analyses concluded that the 7 YY1 binding sites are not required for the establishment of
the allele-specific DNA methylation of the Peg3-DMR during oogenesis, but that some of these YY1 binding sites may be required for protecting DNA methylation during the spermatogenesis.
MUTATIONAL EFFECTS ON TRANSCRIPTIONAL LEVELS We also analyzed the potential effects of the mutation on the transcriptional levels of the genes within the _Peg3_ domain. Total RNA was
isolated from the brains of the two sets of one-day-old pups for cDNA synthesis (Fig. 4). The first set includes the wild-type and heterozygote of both sexes with the paternal transmission
of the mutant allele (Fig. 4A,C), whereas the second set includes the same combination of pups but with the maternal transmission of the mutant allele (Fig. 4B). These two sets of cDNA were
used for performing qRT-PCR with several sets of primers targeting the imprinted genes within the _Peg3_ domain, including _Peg3_, _Usp29_, _Zim1_ (Zinc finger protein 1, imprinted) and
_Zfp264_ (Zinc finger protein 264). The other two remaining genes, _Zim2_ (Zinc finger protein 2, imprinted) and _Zim3_ (Zinc finger protein 3, imprinted), were not included for this
analysis due to the very low expression levels in neonatal brains. This series of expression analyses derived the following results. First, the paternal transmission of the mutant allele
resulted in the down-regulation of both _Peg3_ and _Usp29_, suggesting that the YY1 binding sites may function as a transcriptional activator (Fig. 4A). Interestingly, the levels of the
observed down-regulation were not similar between two sexes: 10% (female) vs 48% (male) for _Peg3_ whereas 17% (female) vs 51% (male) for _Usp29_. The mutant allele also caused a similar
sex-biased outcome in the expression levels of the two adjacent genes, _Zim1_ and _Zfp264_: 350% (female) and 89% (male) for _Zim1_ whereas 29% (female) and 174% (male) for _Zfp264_ (Fig.
4C). Second, the maternal transmission of the mutant allele did not cause any major effects on the expression levels of the surrounding genes (Fig. 4B), which is consistent with the fact
that the maternal allele of the Peg3-DMR harboring the mutated YY1 binding sites are inactive due to DNA methylation. Third, we also analyzed the mutational effects using the total RNA
isolated from placenta (Fig. 4D) and also adult tissues, including the brains and hearts of 2-month-old male and female mice (Supplemental material 4). According to the results, the mutation
resulted in the down-regulation of all the genes tested in placenta: _Peg3_ (63%), _Usp29_ (86%), _Zim1_ (58%) and _Zfp264_ (77%). Among these changes, the expression level changes of
_Peg3_ and _Zim1_ were statistically significant. In the case for adult tissues, both _Peg3_ and _Usp29_ were down-regulated in the brain and heart of adult mice (Supplemental material 4).
Overall, this series of expression analyses concluded that the mutation on YY1 binding sites resulted in the overall down-regulation of _Peg3_ and _Usp29_ in the tissues of neonatal and
adult stages. Interestingly, the levels of the observed down-regulation of _Peg3_ and _Usp29_ differ between males and females in neonatal brains, suggesting that the functional involvement
of YY1 in the _Peg3_ domain might be different between the two sexes. MUTATIONAL EFFECTS ON CHROMATIN STRUCTURE The mutational effects were further analyzed using MEFs (Mouse Embryonic
Fibroblasts) derived from a set of 13.5-d.p.c. embryos that had been prepared through timed mating between male heterozygotes and female breeders (Fig. 5). First, the total RNA isolated from
the male and female sets of MEFs, WT and KO, were used for measuring the expression levels of the genes within the _Peg3_ domain. The result from the male set is shown as a representative
set since the changes observed from two sexes were similar (Fig. 5A,B). The outcome is overall similar to those from neonatal brains: the dramatic down-regulation of _Peg3_ (14%) and _Usp29_
(10%) and the concurrent up-regulation of _Zim1_ (442%). The mutation also resulted in the less dramatic down-regulation of _Zfp264_ (51%). The observed down-regulation of _Peg3_ and
_Usp29_ was consistent with the patterns from neonatal and adult tissues. The observed changes in the expression levels were also analyzed in terms of their DNA methylation levels (Fig. 5C).
The results revealed no major change in the DNA methylation levels of the imprinted genes in both female and male MEFs, suggesting that any changes in the expression levels of the imprinted
genes are likely caused by some unknown factors other than DNA methylation. We further followed up the potential effect of the mutation on the chromatin structure of the Peg3-DMR in the
following manner. The histone modification profiles of the Peg3-DMR are already available as part of the Epigenome project (Fig. 6A). According to the available data, the 2.5-kb YY1 binding
region is represented by two H3K27ac peaks, and these two peaks are also surrounded by several DNaseI-hypersensitive sites, which are marked as dark areas. Interestingly, YY1 binding sites
tend to be located in the border regions between the H3K27ac peaks and DNaseI-hypersensitive sites, suggesting that YY1 may be involved in the formation and/or maintenance of the chromatin
environment of the Peg3-DMR. To test this possibility, we performed two sets of ChIP (Chromatin ImmunoPrecipitation) experiments. First, we tested whether the mutation indeed abrogates the
binding of YY1 to the Peg3-DMR. The chromatin prepared from a set of WT and KO MEF cells were immunoprecipitated with anti-YY1 polyclonal antibody, and the precipitated DNA was analyzed with
qPCR analyses (Fig. 6B). As expected, the levels of the enrichment were much lower in the KO MEF than the levels in the WT MEF at all of the YY1 binding sites, confirming that the mutation
indeed abrogated the binding of YY1 to the Peg3-DMR. Second, a similar series of ChIP experiments were also performed with anti-H3K27ac antibody (Fig. 6B). According to the results, the
three regions showed high levels of the H3K27ac modification in the KO MEF than in the WT MEF, including YY1-01, YY1-34 and YY1-45. In contrast, two regions showed no difference between the
two pools of MEFs, YY1-23 and YY1-56. The higher levels of H3K27ac modification observed from the KO MEF was unexpected since the mutation causing the down-regulation of both _Peg3_ and
_Usp29_ should, intuitively, correlate well with the loss of the activation signals of histone modifications, such as H3K27ac. Although unexpected, the results are overall consistent with
the initial prediction that YY1 may be involved in the formation of the proper chromatin environment of the Peg3-DMR. Taken together, this series of analyses using MEFs concluded that the
mutation resulted in the down-regulation of _Peg3_ and _Usp29_ and also caused the changes in the histone modification levels associated with the Peg3-DMR. DISCUSSION In the current study,
we have mutated the 7 YY1 binding sites localized within the Peg3-DMR. According to the results, the mutation resulted in the down-regulation of the two adjacent genes, _Peg3_ and _Usp29_,
in the majority of the tissues harvested at different developmental stages, suggesting YY1 as an activator for the transcription of the two genes. In contrast, the mutation did not cause any
effect on the allele-specific DNA methylation of the Peg3-DMR, confirming that YY1 is not involved in the DNA methylation of the _Peg3_ domain. Overall, the current study concluded that YY1
is mainly involved in controlling the transcriptional levels of the _Peg3_ domain. The unusual cluster of multiple YY1 binding sites within the Peg3-DMR has been a very unique feature that
has been conserved during mammalian evolution7. Similar clusters of YY1 and CTCF binding sites are also present within the ICRs of the other imprinted domains, including the _Gnas_ and
_H19_/_Igf2_ domains7. Thus, these clusters of YY1 and CTCF binding sites have been thought to be involved in some unknown functions that are unique to genomic imprinting. One such function
may be related to the establishment and maintenance of allele-specific DNA methylation. This prediction has been further supported by CTCF’s roles in protecting DNA methylation from the
active maternal allele of the ICR of the _H19_/_Igf2_ domain9, 11. A series of similar observations have been also made from the YY1 binding sites in the _Peg3_ and _Gnas_ domains, revealing
that reducing the protein levels of YY1 caused either DNA hypomethylation or hypermethylation on the ICRs of the _Peg3_ and _Gnas_ domains13,14,15. Despite all these previous observations,
the current study clearly demonstrated that the YY1 binding sites within the Peg3-DMR are not directly involved in the establishment and maintenance of the allele-specific DNA methylation on
the Peg3-DMR (Fig. 3). This further corrects the previous interpretation of the results that were derived from a series of YY1 knockdown experiments13,14,15. In those experiments, the
changes observed in the DNA methylation levels of the Peg3-DMR may have been an indirect outcome through some unknown loci that are controlled by YY1. On a separate note, however, a subset
of the male heterozygotes was shown to produce sperm with DNA hypermethylation on the Peg3-DMR (Fig. 3). Interestingly, a similar result was also observed from the previous experiments,
displaying DNA hypermethylation on the Peg3-DMR in the sperm that had underwent conditional knockdown of YY1 during spermatogenesis15. Thus, it is still possible that some of the YY1 binding
sites may be involved in protecting the encroachment of DNA methylation from the adjacent regions to the Peg3-DMR. This is reminiscent of the case of the CTCF sites within the ICR of the
_H19_/_Igf2_ domain9, 11. Overall, the current study along with the previous studies suggest that one of the shared functions between YY1 and CTCF in genomic imprinting may be protecting the
active allele of ICRs from DNA methylation. The results from the tissues of neonatal and adult stages clearly demonstrated that both _Peg3_ and _Usp29_ are down-regulated in the mutant
mice, suggesting that the YY1 binding sites function as an activator for the transcription of _Peg3_ and _Usp29_ (Figs 4 and 5). This down-regulation was also accompanied with the changes in
the transcription levels of the adjacent genes within the _Peg3_ domain, including _Zim1_ and _Zfp264_ (Fig. 4). _Zim1_ was up-regulated in the mutants, whereas _Zfp264_ was either up and
down-regulated depending upon the individual tissues tested. These changes observed from _Zim1_ and _Zfp264_ are consistent with the pattern observed from the other ICRs, further supporting
the fact that the Peg3-DMR with the 7 YY1 binding sites is responsible for controlling the transcription of the entire _Peg3_ domain. On the other hand, the current study also provides one
unique observation, which has not been observed from the other ICRs. The levels of the down-regulation of _Peg3_ and _Usp29_ were different between males and females (Fig. 4A). The
mutational effects on females observed in neonatal brains were more severe than those on males, which might be also responsible for the different responses from the two neighboring genes,
_Zim1_ and _Zfp264_. It has also been known that the _Peg3_ locus is sex-specific in many ways. For instance, the expression levels of _Peg3_ itself were different between two sexes, showing
higher in males than in females20. Also, the mutations and environmental stresses affecting the _Peg3_ locus have been shown to affect more severely the survival of males than females21,
22. The current study further suggests that YY1 might be the main culprit causing these sex-biased effects, which will be interesting to pursue in the near future. Overall, the current study
concluded that the main role of the YY1 binding sites within the Peg3-DMR is controlling the transcriptional levels of _Peg3_ and _Usp29_. Despite numerous studies, it is still enigmatic
why the Peg3-DMR and the other ICRs have maintained a cluster of multiple YY1 or CTCF binding sites during mammalian evolution. Two scenarios have been previously proposed for this unusual
multiplicity7. The first scenario is that multiple DNA-binding sites for YY1 or CTCF may be needed for the maintenance of the epigenetic modifications for relatively large-size regulatory
regions, ICRs, which range from 2 to 4 kb in length. The results from the current study support this possibility since the mutation on YY1 binding sites rendered the encroachment of DNA
methylation from the adjacent region to the Peg3-DMR during spermatogenesis (Fig. 3). Also, the available histone modification profiles indicate that the positions of all of the YY1 binding
sites within the Peg3-DMR coincide very well with the border regions between histone modification peaks and DNaseI-hypersensitive regions (Fig. 6A). These lines of evidence clearly support
an idea that the multiple YY1 binding sites are involved in the formation of the chromatin environment of the Peg3-DMR. The second scenario is that the multiple YY1-binding sites within the
Peg3-DMR might be designed to secure a sufficient level of the YY1 protein around this ICR. Yet, this protein level of YY1 around the Peg3-DMR might be a very critical factor deciding the
body size of animal pups. This is likely since the gene dosage of _Peg3_ is known to correlate very well with the body size of animal pups21, 23. In this case, the increased levels of the
YY1 protein through additional YY1 binding sites might boost the body size and thus the survival chance of animals. On the other hand, the unusually high levels of the YY1 protein could
cause the unsustainable size of the fetus, which might jeopardize the health of the pregnant females. To counter this hazardous outcome, the maternal genome might need to reduce the number
of YY1 binding sites through DNA methylation-mediated substitutions from CpG to TpG or CpA within the YY1-binding site (CGCCATnTT), which has been shown to reduce dramatically the binding
affinity of YY1 to DNA24. Consistent with this, the Peg3-DMRs of many mammals, including human and mouse, have several imperfect YY1 binding sites with one or two base differences from the
consensus sequence. This decay process of CpG sites during evolution is particularly feasible since the Peg3-DMR becomes methylated as part of genomic imprinting during oogenesis. If this
scenario turns out to be the case, the multiple YY1 binding sites detected in a given species might be simply reflecting the outcome of this evolutionary tug-of-war between two parental
genomes with opposite needs25, 26. Further investigation of this possibility would be of great interest, but may require different experimental approaches involving phylogenetic and
comparative genomics. Overall, the observations from the current study are consistent with both scenarios, which are non-exclusive. Therefore, we conclude that the multiple YY1 binding sites
within the Peg3-DMR may have been selected for both functional needs, maintaining the chromatin environment and securing a sufficient level of the YY1 protein around the Peg3-DMR. MATERIALS
AND METHODS ETHICS STATEMENT All the experiments related to mice were performed in accordance with National Institutes of Health guidelines for care and use of animals, and also approved by
the Louisiana State University Institutional Animal Care and Use Committee (IACUC), protocol #16-060. GENERATION OF A KNOCKOUT ALLELE The targeting vector for the knockout experiment was
constructed with the RED/ET recombination technique (Gene Bridges; ref. 27). A mouse BAC (bacterial artificial chromosome) clone, RP23-178C5 (Invitrogen), was used as a source for isolating
the 15.4-kb genomic fragment surrounding the Peg3-DMR (nucleotide positions 6,718,442-6,733,840 in the mouse chromosome 7 of mm10). The 15.4-kb fragment was isolated through homologous
recombination using two hooks, which were part of the two following oligonucleotides: GB primer 1 (5′-GCAAACGCCGTGTTATCAAACACCTTCATCTCAGACCACGGTCTGTGCTG-_GTCGAC-_ACAGCTTGTCTGTAAGCGGATG-3′)
and GB primer 2 (5′-CAAAACAGACAACTGTGAAAAACTCACCACTCCGTTGGAGAGTTTCAAGA- _GCGGCCGC-_GCTCTCCTGAGTAGGACAAATCCG-3′). The 50 nucleotide-long sequences at the 5′-ends of both primers were the
homology hooks while the sequences at the 3′-end of both primers were included for the amplification of a minimal cassette for the RED/ET recombination system (Gene Bridges, Cat. No. K002).
Two restriction enzyme sites, _Sal_I and _Not_I (italicized regions), were also included as part of the sequences to be used for the linearization of the final vector and the cloning of the
negative selection marker DTA (Diphtheria toxin A), respectively. PCR amplification of the minimal cassette with these two primers generated the linearized minimal cassette with the two
homology hooks at its 5′- and 3′-ends. The E. coli strain carrying the BAC RP23-178C5 was transformed with the expression plasmid pRedET and the linearized minimal cassette containing the
two homology hooks. Several colonies containing the circularized 17.8-kb vector (15.4-kb target fragment plus the 2.4-kb minimal cassette) were obtained through the ampicillin selection. The
integrity of the isolated 15.4-kb genomic fragment was further confirmed through a series of restriction enzyme digestions. One round of additional pRedET-based recombination was performed
to replace the 2.5-kb YY1 binding region with the neomycin resistance gene (_NeoR_) that is flanked by FRT sites. This replacement process has also incorporated one loxP site at the 5′-end
and one _BamH_I site at the 3′-end (Fig. 2A). The subsequent vector was further modified by inserting the mutant version of the YY1 binding region into the _BamH_I site. The mutant version
of the YY1-binding region, which was previously generated16, was amplified with the two primers containing _BamH_I site for subcloning. During this subcloning step, another loxP site has
been inserted at the 3′-side, thus this newly inserted loxP site can be paired with the other loxP site that had been inserted earlier at the 5′-side of the 2.5-kb YY1 binding region.
Finally, we subcloned the expression cassette DTA into the _Not_I site as a negative selection marker. The final 21.4-kb KO vector was linearized with _Sal_I digestion, and subsequently used
for transfection into the AB2.2 ES cell line of the 129/SvJ origin (http://www.bcm.edu/dtmc/, Darwin Transgenic Mouse Core facility of Baylor College of Medicine). Transfected ES cells were
first screened with a long-distance PCR scheme that can confirm the proper recombination of the 5′-side genomic fragment with the following primer set: Peg3-KO-65
(5′-TTCCTAAAGGCAAGTAGGACCT-3′) and Neo-R1 (5′-GATTCGCAGCGCATCGCCTTCT-3′). Later, a subset of the potential targeted clones identified through LD-PCR were further analyzed with southern
blotting to confirm the proper recombination of the 3′-side genomic fragment. For this southern blotting, the DNA isolated from ES cells were digested with _BamH_I, and probed with the
495-bp fragment that had been amplified by PCR with the following primer set: Peg3-KO-63 (5′-ACCTTCCACTAGATTTCACCTCCT-3′) and Peg3-KO-64 (5′-CACTGCCAAAAGCATGAGATGGTC-3′). Two targeted ES
cell was microinjected into the blastocysts of e3.5-embryos of the C57BL/6 J (B6) mouse, generating ten chimeras with varying degrees of coat color contribution. Four of these chimeras were
bred with 8 B6 females, finally deriving F1 mice with germline transmission of the targeted allele. MOUSE BREEDING The male and female heterozygotes carrying the mutant allele were bred
individually with female and male wild-type littermates. One-day-old pups derived from these breeding experiments were analyzed in terms of sex, genotype and body weight. Statistical
significance of potential difference of litter size and average weight between two breeding experiments was tested Χ2-test. All the mice were housed at the DLAM (Division of Lab Animal
Medicine) of LSU on a regular 12-12 dark-light cycle under a constant temperature 70°F and 50% humidity. All animals were given ad libitum access to water and Rodent Diet 5001. The nursing
females were with Mouse Diet 5015. The mice were euthanized by CO2 asphixation in accordance with the rules and regulations set forth by the IACUC. For genotyping, genomic DNA was isolated
from either clipped ears by incubating the tissues overnight at 55 °C in the lysis buffer (0.1 M Tris-Cl, pH 8.8, 5 mM EDTA, pH 8.0, 0.2% SDS, 0.2 M NaCl, 20 μg/ml Proteinase K). The
isolated DNA was subsequently genotyped using the following primer set: KO-type-F (5′-ATGACAAGTGGGCTTGCTGCAG-3′) and KO-type-R (5′-GGATGTAAGATGGAGGCACTGT-3′). The sexes of the pups were
determined through PCR using the following primer set: mSry-F (5′-GTCCCGTGGTGAGAGGCACAAG-3′) and mSry-R (5′-GCAGCTCTACTCCAGTCTTGCC-3′). EXPRESSION ANALYSES AND IMPRINTING TEST Total RNA was
isolated from the tissues of one-day-old heads and adult mice using a commercial kit (Trizol, Invitrogen). The total RNA was then reverse-transcribed using the M-MuLV kit (Invitrogen), and
the subsequent cDNA was used as a template for quantitative real-time PCR. This analysis was performed with the iQ SYBR green supermix (Bio-Rad) using the ViiA™ 7 Real-Time PCR System (Life
Technologies). All qRT-PCR reactions were carried out for 40 cycles under standard PCR conditions. The analyses of the results derived from qRT-PCR were described previously13. Statistical
significance of potential difference of expression levels of a given gene between two samples was tested Student’s t-test. The information regarding individual primer sequences and PCR
conditions has been published in the previous study21. For imprinting test, the heterozygotes of the 129/B6 background were bred with the PWD/PhJ strain (Jackson Lab, Stock No. 004660). The
F1 hybrid of this crossing was used for isolating total RNA. The polymorphisms and restriction enzymes used for each gene’s imprinting test are also available through the previous study21.
DNA METHYLATION ANALYSIS DNA methylation levels of each target region were analyzed using genomic DNA isolated from two germ cells, sperm and oocyte, and also the tissues of the neonates and
adult mice. Detailed protocols for isolating sperm and oocyte were described previously15. Briefly, sperm was isolated from the epididymus of 3 and 6-moth-old mice using the ‘swim-up’
method28. Mature oocytes were isolated from 3-month-old females after superovulation with PMSG and hCG treatment29, 30. The isolated DNA was treated with the bisulfite conversion reaction
according to the manufacturer’s protocol (EZ DNA methylation kit, Zymo Research). The converted DNA was used as a template for the PCR reaction using specific primers that were designed for
amplifying each target region. The majority of PCR amplifications were carried out using a standard protocol with 40 cycles, whereas the PCR amplification from eggs was conducted using a
nested scheme with the initial step with 40 cycles followed by the second step with 30 cycles. Each PCR product was further analyzed using the following two approaches: 1) the restriction
enzyme digestion-based COBRA19 and 2) subcloning and sequencing. For the COBRA analysis, each PCR product was digested with a series of restriction enzymes. The PCR product was also
individually subcloned into the pGEM T-Easy vector (Promega), and 10 to 20 clones were subsequently sequenced to survey its DNA methylation levels at each locus. The detailed information
regarding oligonucleotide sequences, sequence polymorphisms, and COBRA is also available (Supplemental material 5). DERIVATION OF MEF (MOUSE EMBRYONIC FIBROBLAST) CELLS Two litters of
13.5-dpc embryos of the 129/B6 background were harvested through timed mating of the male KO heterozygotes with the female wild-type littermates. The head portion and the red tissues were
removed from the embryos, and the remaining portions were minced with razor blades. These minced tissues were transferred to a 15 ml conical tubes containing 1 ml trypsin (Invitrogen, Cat.
No. 25300062). After 5 min incubation at 37 °C, the cells were harvested with centrifugation, and later resuspended in 15 ml media (Life technologies, Cat. No.10566024). Finally, the
resuspended cells were plated onto a T-75 flask. The MEF from each embryo was first genotyped using the following primer set: Peg3-KO-63 (5′-ACCTTCCACTAGATTTCACCTCCT-3′) and Peg3-KO-64
(5′-CACTGCCAAAAGCATGAGATGGTC-3′). The sex of each MEF was also determined using the following primer set: mSry-F (5′-GTCCCGTGGTGAGAGGCACAAG-3′) and mSry-R (5′-GCAGCTCTACTCCAGTCTTGCC-3′).
CHROMATIN IMMUNOPRECIPITATION (CHIP) Chromatins were prepared from MEF according to the method previously described5. In brief, the homogenized samples were first cross-linked with 1%
formaldehyde for 20 mins, and then lysed with the buffer containing protease inhibitor cocktail (Millipore, Cat. No. 539131). The released nuclei were fractionated with sonication to derive
a pool of DNA fragments size-ranging from 300 to 500 bp in length. The prepared chromatin was immunoprecipitated with two commercial antibodies: anti-YY1 antibody (SantaCruz Biotech, Cat.
No. sc-1703) and anti-H3K27ac antibody (Abcam, Cat. No. ab4729). The immunoprecipitated DNA was dissolved in 80 μl of TE for PCR analyses. REFERENCES * Barlow, D. P. & Bartolomei, M. S.
Genomic imprinting in mammals. _Cold Spring Harb Perspect Biol_ 6(2), doi:10.1101/cshperspect.a018382 (2014). * Bartolomei, M. S. & Ferguson-Smith, A. C. Mammalian genomic imprinting.
_Cold Spring Harb Perspect Biol_ 3(7), doi:10.1101/cshperspect.a002592 (2011). * Bell, A. C. & Felsenfeld, G. Methylation of a CTCF-dependent boundary controls imprinted expression of
the Igf2 gene. _Nature_ 405, 482–485, doi:10.1038/35013100 (2000). Article ADS CAS PubMed Google Scholar * Hark, A. T. _et al_. CTCF mediates methylation-sensitive enhancer-blocking
activity at the H19/Igf2 locus. _Nature_ 405, 486–489, doi:10.1038/35013106 (2000). Article ADS CAS PubMed Google Scholar * Kim, J., Kollhoff, A., Bergmann, A. & Stubbs, L.
Methylation-sensitive binding of transcription factor YY1 to an insulator sequence within the paternally expressed imprinted gene, Peg3. _Hum Mol Genet_ 12, 233–245 (2003). Article CAS
PubMed Google Scholar * Kim, J. D. _et al_. Identification of clustered YY1 binding sites in imprinting control regions. _Genome Res_ 16, 901–911, doi:10.1101/gr.5091406 (2006). Article
CAS PubMed PubMed Central Google Scholar * Kim, J. Multiple YY1 and CTCF binding sites in imprinting control regions. _Epigenetics_ 3, 115–118 (2008). Article PubMed Google Scholar *
Leighton, P. A., Saam, J. R., Ingram, R. S., Stewart, C. L. & Tilghman, S. M. An enhancer deletion affects both H19 and Igf2 expression. _Genes Dev_ 9, 2079–2089 (1995). Article CAS
PubMed Google Scholar * Schoenherr, C. J., Levorse, J. M. & Tilghman, S. M. CTCF maintains differential methylation at the Igf2/H19 locus. _Nat Genet_ 33, 66–69, doi:10.1038/ng1057
(2003). Article CAS PubMed Google Scholar * Engel, N., West, A. G. & Felsenfeld, G. Bartolomei MS. Antagonism between DNA hypermethylation and enhancer-blocking activity at the H19
DMD is uncovered by CpG mutations. _Nat Genet_ 36, 883–888, doi:10.1038/ng1399 (2004). Article CAS PubMed Google Scholar * Fedoriw, A. M., Stein, P., Svoboda, P., Schultz, R. M. &
Bartolomei, M. S. Transgenic RNAi reveals essential function for CTCF in H19 gene imprinting. _Science_ 303, 238–240, doi:10.1126/science.1090934 (2004). Article ADS CAS PubMed Google
Scholar * Enge, lN., Thorvaldsen, J. L. & Bartolomei, M. S. CTCF binding sites promote transcription initiation and prevent DNA methylation on the maternal allele at the imprinted
H19/Igf2 locus. _Hum Mol Genet_ 15, 2945–2954, doi:10.1093/hmg/ddl237 (2006). Article Google Scholar * Kim, J. D., Hinz, A. K., Choo, J. H., Stubbs, L. & Kim, J. YY1 as a controlling
factor for the Peg3 and Gnas imprinted domains. _Genomics_ 89, 262–269, doi:10.1016/j.ygeno.2006.09.009 (2007). Article CAS PubMed Google Scholar * Kim., J. & Kim, J. D. _In vivo_
YY1 knockdown effects on genomic imprinting. _Hum Mol Genet_ 17, 391–401, doi:10.1093/hmg/ddm316 (2008). Article CAS PubMed Google Scholar * Kim, J. D., Kang, K. & Kim, J. YY1’s role
in DNA methylation of Peg3 and Xist. _Nucleic Acids Res_ 37, 5656–5664, doi:10.1093/nar/gkp613 (2009). Article CAS PubMed PubMed Central Google Scholar * Kim, J. D., Yu, S., Choo, J.
H. & Kim, J. Two evolutionarily conserved sequence elements for Peg3/Usp29 transcription. _BMC Mol Biol_ 9, 108, doi:10.1186/1471-2199-9-108 (2008). Article PubMed PubMed Central
Google Scholar * Perera, B. P., Teruyama, R. & Kim, J. Yy1 gene dosage effect and bi-allelic expression of Peg3. _PLoS One_ 10, e0119493, doi:10.1371/journal.pone.0119493 (2015).
Article PubMed PubMed Central Google Scholar * Kim, J. D. & Kim, J. YY1’s longer DNA-binding motifs. _Genomics_ 93, 152–158, doi:10.1016/j.ygeno.2008.09.013 (2009). Article CAS
PubMed Google Scholar * Xiong, Z. & Laird, P. W. COBRA: a sensitive and quantitative DNA methylation assay. _Nucleic Acids Res_ 25, 2532–2534 (1997). Article CAS PubMed PubMed
Central Google Scholar * Faisal, M., Kim, H. & Kim, J. Sexual differences of imprinted genes’ expression levels. _Gene_ 533, 434–438, doi:10.1016/j.gene.2013.10.006 (2014). Article
CAS PubMed Google Scholar * Kim, J. _et al_. Imprinting control region (ICR) of the Peg3 domain. _Hum Mol Genet_ 21, 2677–2687, doi:10.1093/hmg/dds092 (2012). Article CAS PubMed PubMed
Central Google Scholar * Radford, E. J. _et al_. An unbiased assessment of the role of imprinted genes in an intergenerational model of developmental programming. _PLoS Genet_ 8,
e1002605, doi:10.1371/journal.pgen.1002605 (2012). Article CAS PubMed PubMed Central Google Scholar * Kim, J. _et al_. Peg3 mutational effects on reproduction and placenta-specific gene
families. _PLoS One_ 8, e83359, doi:10.1371/journal.pone.0083359 (2013). Article ADS PubMed PubMed Central Google Scholar * Kim, J. D., Faulk, C. & Kim, J. Retroposition and
evolution of the DNA-binding motifs of YY1, YY2 and REX1. _Nucleic Acids Res_ 35, 3442–3452, doi:10.1093/nar/gkm235 (2007). Article CAS PubMed PubMed Central Google Scholar * Moore, T.
& Haig, D. Genomic imprinting in mammalian development: a parental tug-of-war. _Trends Genet_ 7, 45–49, doi:10.1016/0168-9525(91)90230-N (1991). Article CAS PubMed Google Scholar *
Haig, D. Genomic imprinting and kinship: how good is the evidence? _Annu Rev Genet_ 38, 553–585, doi:10.1146/annurev.genet.37.110801.142741 (2004). Article CAS PubMed Google Scholar *
Angrand, P. O., Daigle, N., van der Hoeven, F., Scholer, H. R. & Stewart, A. F. Simplified generation of targeting constructs using ET recombination. _Nucleic Acids Res_ 27, e16 (1999).
Article CAS PubMed PubMed Central Google Scholar * Bunch, D. O. & Saling, P. M. Generation of a mouse sperm membrane fraction with zona receptor activity. _Biol Reprod_ 44, 672–680
(1991). Article CAS PubMed Google Scholar * Eppig, J. J. & Telfer, E. E. Isolation and culture of oocytes. _Methods Enzymol_ 225, 77–84 (1993). Article CAS PubMed Google Scholar
* Hogan, B., Beddington, R., Costantini, F. & Lacy, E. _Manipulating the mouse embryo: A laboratory Manual, 2_ _nd_ _ed_. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
(1994). Download references ACKNOWLEDGEMENTS We would like to thank Drs Hana Kim and Michelle Thiaville for their help in the initial stage of the construction of KO vectors. We also like to
thank the members of JooKim Lab for their thoughtful feedback and discussion on the manuscript. This research was supported by the National Institute of Health (R01-GM066225 and
R01-GM097074 to J.K.). AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Department of Biological Sciences, Louisiana State University, Baton Rouge, LA, 70803, USA Hongzhi He, An Ye,
Bambarendage P. U. Perera & Joomyeong Kim Authors * Hongzhi He View author publications You can also search for this author inPubMed Google Scholar * An Ye View author publications You
can also search for this author inPubMed Google Scholar * Bambarendage P. U. Perera View author publications You can also search for this author inPubMed Google Scholar * Joomyeong Kim View
author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS H.H. and J.K. conceived the experiments and wrote the manuscript; H.H., B.P. and A.Y. conducted
the experiments; J.K., H.H., B.P. and A.Y. analyzed the results. All authors reviewed the manuscript. CORRESPONDING AUTHOR Correspondence to Joomyeong Kim. ETHICS DECLARATIONS COMPETING
INTERESTS The authors declare that they have no competing interests. ADDITIONAL INFORMATION PUBLISHER'S NOTE: Springer Nature remains neutral with regard to jurisdictional claims in
published maps and institutional affiliations. ELECTRONIC SUPPLEMENTARY MATERIAL SUPPLEMENTARY INFORMATION RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative
Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the
original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in
the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended
use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit
http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE He, H., Ye, A., Perera, B.P.U. _et al._ YY1’s role in the _Peg3_ imprinted domain.
_Sci Rep_ 7, 6427 (2017). https://doi.org/10.1038/s41598-017-06817-5 Download citation * Received: 13 January 2017 * Accepted: 19 June 2017 * Published: 25 July 2017 * DOI:
https://doi.org/10.1038/s41598-017-06817-5 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not
currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative