Mechanism of autocatalytic activation during proteasome assembly
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT Many large molecular machines are too elaborate to assemble spontaneously and are built through ordered pathways orchestrated by dedicated chaperones. During assembly of the core
particle (CP) of the proteasome, where protein degradation occurs, its six active sites are simultaneously activated via cleavage of N-terminal propeptides. Such activation is autocatalytic
and coupled to fusion of two half-CP intermediates, which protects cells by preventing activation until enclosure of the active sites within the CP interior. Here we uncover key mechanistic
aspects of autocatalytic activation, which proceeds through alignment of the β5 and β2 catalytic triad residues, respectively, with these triads being misaligned before fusion. This
mechanism contrasts with most other zymogens, in which catalytic centers are preformed. Our data also clarify the mechanism by which individual subunits can be added in a precise, temporally
ordered manner. This work informs two decades-old mysteries in the proteasome field, with broader implications for protease biology and multisubunit complex assembly. Access through your
institution Buy or subscribe This is a preview of subscription content, access via your institution ACCESS OPTIONS Access through your institution Access Nature and 54 other Nature Portfolio
journals Get Nature+, our best-value online-access subscription $32.99 / 30 days cancel any time Learn more Subscribe to this journal Receive 12 print issues and online access $209.00 per
year only $17.42 per issue Learn more Buy this article * Purchase on SpringerLink * Instant access to full article PDF Buy now Prices may be subject to local taxes which are calculated
during checkout ADDITIONAL ACCESS OPTIONS: * Log in * Learn about institutional subscriptions * Read our FAQs * Contact customer support SIMILAR CONTENT BEING VIEWED BY OTHERS VISUALIZING
CHAPERONE-MEDIATED MULTISTEP ASSEMBLY OF THE HUMAN 20S PROTEASOME Article Open access 10 April 2024 STRUCTURE OF THE PREHOLOPROTEASOME REVEALS LATE STEPS IN PROTEASOME CORE PARTICLE
BIOGENESIS Article 31 August 2023 STRUCTURES OF CHAPERONE-ASSOCIATED ASSEMBLY INTERMEDIATES REVEAL COORDINATED MECHANISMS OF PROTEASOME BIOGENESIS Article 12 April 2021 DATA AVAILABILITY
Cryo-EM maps (EMD-41963, EMD-41993) and atomic model coordinates (PDB 8U6Y, PDB 8U7U) have been deposited in the EMDB and RCSB, respectively. Other PDB structures used here include 5CZ4,
7LS6 and 8T08. Source data are provided with this paper. REFERENCES * Jäger, S., Groll, M., Huber, R., Wolf, D. H. & Heinemeyer, W. Proteasome beta-type subunits: unequal roles of
propeptides in core particle maturation and a hierarchy of active site function. _J. Mol. Biol._ 291, 997–1013 (1999). Article PubMed Google Scholar * Schnell, H. M., Walsh, R. M.,
Rawson, S. & Hanna, J. Chaperone-mediated assembly of the proteasome core particle – recent developments and structural insights. _J. Cell Sci._ 135, jcs259622 (2022). Article CAS
PubMed PubMed Central Google Scholar * Watanabe, A., Yashiroda, H., Ishihara, S., Lo, M. & Murata, S. The molecular mechanisms governing the assembly of the immuno- and
thymoproteasomes in the presence of constitutive proteasomes. _Cells_ 11, 1580 (2022). Article CAS PubMed PubMed Central Google Scholar * Chen, P. & Hochstrasser, M. Autocatalytic
subunit processing couples active site formation in the 20S proteasome to completion of assembly. _Cell_ 86, 961–972 (1996). Article CAS PubMed Google Scholar * Hirano, Y. et al.
Dissecting beta-ring assembly pathway of the mammalian 20S proteasome. _EMBO J._ 27, 2204–2213 (2008). Article CAS PubMed PubMed Central Google Scholar * Schnell, H. M. et al.
Structures of chaperone-associated assembly intermediates reveal coordinated mechanisms of proteasome biogenesis. _Nat. Struct. Mol. Biol._ 28, 418–425 (2021). Article CAS PubMed PubMed
Central Google Scholar * Seemüller, E., Lupas, A. & Baumeister, W. Autocatalytic processing of the 20S proteasome. _Nature_ 382, 468–470 (1996). Article PubMed Google Scholar *
Huber, E. M. et al. A unified mechanism for proteolysis and autocatalytic activation in the 20S proteasome. _Nat. Commun._ 7, 10900 (2016). Article CAS PubMed PubMed Central Google
Scholar * Groll, M. et al. Structure of 20S proteasome from yeast at 2.4 Å resolution. _Nature_ 386, 463–471 (1997). Article CAS PubMed Google Scholar * Ramos, P. C., Marques, A. J.,
London, M. K. & Dohmen, R. J. Role of C-terminal extensions of subunits beta2 and beta7 in assembly and activity of eukaryotic proteasomes. _J. Biol. Chem._ 279, 14323–14330 (2004).
Article CAS PubMed Google Scholar * Walsh, R. M. et al. Structure of the preholoproteasome reveals late steps in proteasome core particle biogenesis. _Nat. Struct. Mol. Biol._ 30,
1516–1524 (2023). Article PubMed PubMed Central Google Scholar * Gerlinger, U. M., Gückel, R., Hoffmann, M., Wolf, D. H. & Hilt, W. Yeast cycloheximide-resistant crl mutants are
proteasome mutants defective in protein degradation. _Mol. Biol. Cell_ 8, 2487–2499 (1997). Article CAS PubMed PubMed Central Google Scholar * Kock, M. et al. Proteasome assembly from
15S precursors involves major conformational changes and recycling of the Pba1–Pba2 chaperone. _Nat. Commun._ 6, 6123 (2015). Article CAS PubMed Google Scholar * Li, X., Li, Y., Arendt,
C. S. & Hochstrasser, M. Distinct elements in the proteasomal β5 subunit propeptide required for autocatalytic processing and proteasome assembly. _J. Biol. Chem._ 291, 1991–2003 (2016).
Article CAS PubMed Google Scholar * Xie, Y. & Varshavsky, A. RPN4 is a ligand, substrate, and transcriptional regulator of the 26S proteasome: a negative feedback circuit. _Proc.
Natl Acad. Sci. USA_ 98, 3056–3061 (2001). Article CAS PubMed PubMed Central Google Scholar * Guerra-Moreno, A. & Hanna, J. Induction of proteotoxic stress by the mycotoxin patulin.
_Toxicol. Lett._ 276, 85–91 (2017). Article CAS PubMed PubMed Central Google Scholar * Ramos, P. C., Höckendorff, J., Johnson, E. S., Varshavsky, A. & Dohmen, R. J. Ump1p is
required for proper maturation of the 20S proteasome and becomes its substrate upon completion of the assembly. _Cell_ 92, 489–499 (1998). Article CAS PubMed Google Scholar * Marques, A.
J., Glanemann, C., Ramos, P. C. & Dohmen, R. J. The C-terminal extension of the beta7 subunit and activator complexes stabilize nascent 20S proteasomes and promote their maturation. _J.
Biol. Chem._ 282, 34869–34876 (2007). Article CAS PubMed Google Scholar * Arendt, C. S. & Hochstrasser, M. Eukaryotic 20S proteasome catalytic subunit propeptides prevent active
site inactivation by N-terminal acetylation and promote particle assembly. _EMBO J._ 18, 3575–3585 (1999). Article CAS PubMed PubMed Central Google Scholar * Matias, A. C., Matos, J.,
Dohmen, R. J. & Ramos, P. C. Hsp70 and Hsp110 chaperones promote early steps of proteasome assembly. _Biomolecules_ 13, 11 (2022). Article PubMed PubMed Central Google Scholar *
Khan, A. R. & James, M. N. Molecular mechanisms for the conversion of zymogens to active proteolytic enzymes. _Protein Sci._ 7, 815–836 (1998). Article CAS PubMed PubMed Central
Google Scholar * Richter, C., Tanaka, T. & Yada, R. Y. Mechanism of activation of the gastric aspartic proteinases: pepsinogen, progastricsin and prochymosin. _Biochem. J._ 335, 481–490
(1998). Article CAS PubMed PubMed Central Google Scholar * Arolas, J. L., Goulas, T., Cuppari, A. & Gomis-Rüth, F. X. Multiple architectures and mechanisms of latency in
metallopeptidase zymogens. _Chem. Rev._ 118, 5581–5597 (2018). Article CAS PubMed Google Scholar * Poli, M. C. et al. Heterozygous truncating variants in POMP escape nonsense-mediated
decay and cause a unique immune dysregulatory syndrome. _Am. J. Hum. Genet._ 102, 1126–1142 (2018). Article CAS PubMed PubMed Central Google Scholar * de Jesus, A. A. et al. Novel
proteasome assembly chaperone mutations in PSMG2/PAC2 cause the autoinflammatory interferonopathy CANDLE/PRAAS4. _J. Allergy Clin. Immunol._ 143, 1939–1943.e8 (2019). Article PubMed PubMed
Central Google Scholar * Dahlqvist, J. et al. A single-nucleotide deletion in the POMP 5′ UTR causes a transcriptional switch and altered epidermal proteasome distribution in KLICK
genodermatosis. _Am. J. Hum. Genet._ 86, 596–603 (2010). Article CAS PubMed PubMed Central Google Scholar * Ansar, M. et al. Biallelic variants in PSMB1 encoding the proteasome subunit
β6 cause impairment of proteasome function, microcephaly, intellectual disability, developmental delay and short stature. _Hum. Mol. Genet._ 29, 1132–1143 (2020). Article CAS PubMed
Google Scholar * Hwang, G.-W., Ishida, Y. & Naganuma, A. Identification of F-box proteins that are involved in resistance to methylmercury in _Saccharomyces cerevisiae_. _FEBS Lett._
580, 6813–6818 (2006). Article CAS PubMed Google Scholar * Wani, P. S., Rowland, M. A., Ondracek, A., Deeds, E. J. & Roelofs, J. Maturation of the proteasome core particle induces an
affinity switch that controls regulatory particle association. _Nat. Commun._ 6, 6384 (2015). Article CAS PubMed Google Scholar * Weisshaar, N., Welsch, H., Guerra-Moreno, A. &
Hanna, J. Phospholipase Lpl1 links lipid droplet function with quality control protein degradation. _Mol. Biol. Cell_ 28, 716–725 (2017). Article CAS PubMed PubMed Central Google Scholar
* Mastronarde, D. N. Automated electron microscope tomography using robust prediction of specimen movements. _J. Struct. Biol._ 152, 36–51 (2005). Article PubMed Google Scholar * Zheng,
S. Q. et al. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. _Nat. Methods_ 14, 331–332 (2017). Article CAS PubMed PubMed Central Google
Scholar * Rohou, A. & Grigorieff, N. CTFFIND4: fast and accurate defocus estimation from electron micrographs. _J. Struct. Biol._ 192, 216–221 (2015). Article PubMed PubMed Central
Google Scholar * Wagner, T. et al. SPHIRE-crYOLO is a fast and accurate fully automated particle picker for cryo-EM. _Commun. Biol._ 2, 218 (2019). Article PubMed PubMed Central Google
Scholar * Scheres, S. H. W. RELION: implementation of a Bayesian approach to cryo-EM structure determination. _J. Struct. Biol._ 180, 519–530 (2012). Article CAS PubMed PubMed Central
Google Scholar * Punjani, A., Rubinstein, J. L., Fleet, D. J. & Brubaker, M. A. CryoSPARC: algorithms for rapid unsupervised cryo-EM structure determination. _Nat. Methods_ 14, 290–296
(2017). Article CAS PubMed Google Scholar * Zhong, E. D., Bepler, T., Berger, B. & Davis, J. H. CryoDRGN: reconstruction of heterogeneous cryo-EM structures using neural networks.
_Nat. Methods_ 18, 176–185 (2021). Article CAS PubMed PubMed Central Google Scholar * Morin, A. et al. Collaboration gets the most out of software. _eLife_ 2, e01456 (2013). Article
PubMed PubMed Central Google Scholar * Jamali, K. et al. Automated model building and protein identification in cryo-EM maps. _Nature_ https://doi.org/10.1038/s41586-024-07215-4 (2024). *
Pettersen, E. F. et al. UCSF Chimera – a visualization system for exploratory research and analysis. _J. Comput. Chem._ 25, 1605–1612 (2004). Article CAS PubMed Google Scholar * Emsley,
P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. _Acta Crystallogr. D Biol. Crystallogr._ 66, 486–501 (2010). Article CAS PubMed PubMed Central Google
Scholar * Croll, T. I. ISOLDE: a physically realistic environment for model building into low-resolution electron-density maps. _Acta Crystallogr. D Struct. Biol._ 74, 519–530 (2018).
Article CAS PubMed PubMed Central Google Scholar * Liebschner, D. et al. Macromolecular structure determination using X-rays, neutrons and electrons: recent developments in Phenix.
_Acta Crystallogr. D Struct. Biol._ 75, 861–877 (2019). Article CAS PubMed PubMed Central Google Scholar * Kao, A. et al. Development of a novel cross-linking strategy for fast and
accurate identification of cross-linked peptides of protein complexes. _Mol. Cell Proteomics_ 10, M110.002212 (2011). Article PubMed Google Scholar * Gutierrez, C. B. et al. Developing an
acidic residue reactive and sulfoxide-containing MS-cleavable homobifunctional cross-linker for probing protein–protein interactions. _Anal. Chem._ 88, 8315–8322 (2016). Article CAS
PubMed PubMed Central Google Scholar * Wiśniewski, J. R., Zougman, A., Nagaraj, N. & Mann, M. Universal sample preparation method for proteome analysis. _Nat. Methods_ 6, 359–362
(2009). Article PubMed Google Scholar * Jiao, F. et al. Two-dimensional fractionation method for proteome-wide cross-linking mass spectrometry analysis. _Anal. Chem._ 94, 4236–4242
(2022). Article CAS PubMed PubMed Central Google Scholar * Jiao, F. et al. Exploring an alternative cysteine-reactive chemistry to enable proteome-wide PPI analysis by cross-linking
mass spectrometry. _Anal. Chem._ 95, 2532–2539 (2023). Article CAS PubMed PubMed Central Google Scholar * Finley, D., Ozkaynak, E. & Varshavsky, A. The yeast polyubiquitin gene is
essential for resistance to high temperatures, starvation, and other stresses. _Cell_ 48, 1035–1046 (1987). Article CAS PubMed Google Scholar * Gietz, R. D. & Sugino, A. New
yeast–_Escherichia coli_ shuttle vectors constructed with in vitro mutagenized yeast genes lacking six-base pair restriction sites. _Gene_ 74, 527–534 (1988). Article CAS PubMed Google
Scholar Download references ACKNOWLEDGEMENTS We thank J. Roelofs (University of Kansas Medical Center) for the Pba1/2 antibody; H. Schnell for early contributions to the project; and B.
Schulman, F. Adolf, J. Du, E. Goodall, J. W. Harper and D. Finley for helpful discussions. This work was supported by National Institutes of Health grants R01-GM144367 and R01-GM135337 (to
J.H.), and R35-GM145249 (to L.H.). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. AUTHOR INFORMATION Author
notes * These authors contributed equally: Benjamin Velez, Richard M. Walsh Jr, Shaun Rawson. AUTHORS AND AFFILIATIONS * Department of Pathology, Harvard Medical School and Brigham and
Women’s Hospital, Boston, MA, USA Benjamin Velez, Aida Razi, Lea Adams, Erignacio Fermin Perez, Marie Blickling, Tamayanthi Rajakumar, Darlene Fung & John Hanna * Harvard Cryo-Electron
Microscopy Center for Structural Biology, Harvard Medical School, Boston, MA, USA Richard M. Walsh Jr. & Shaun Rawson * Department of Biological Chemistry and Molecular Pharmacology,
Blavatnik Institute, Harvard Medical School, Boston, MA, USA Richard M. Walsh Jr. & Shaun Rawson * Department of Physiology and Biophysics, University of California-Irvine, Irvine, CA,
USA Fenglong Jiao & Lan Huang Authors * Benjamin Velez View author publications You can also search for this author inPubMed Google Scholar * Richard M. Walsh Jr. View author
publications You can also search for this author inPubMed Google Scholar * Shaun Rawson View author publications You can also search for this author inPubMed Google Scholar * Aida Razi View
author publications You can also search for this author inPubMed Google Scholar * Lea Adams View author publications You can also search for this author inPubMed Google Scholar * Erignacio
Fermin Perez View author publications You can also search for this author inPubMed Google Scholar * Fenglong Jiao View author publications You can also search for this author inPubMed Google
Scholar * Marie Blickling View author publications You can also search for this author inPubMed Google Scholar * Tamayanthi Rajakumar View author publications You can also search for this
author inPubMed Google Scholar * Darlene Fung View author publications You can also search for this author inPubMed Google Scholar * Lan Huang View author publications You can also search
for this author inPubMed Google Scholar * John Hanna View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS B.V., J.H., L.A., E.F.P., M.B., A.R.,
T.R. and D.F. performed the biochemical aspects of the work. R.M.W. and S.R. performed cryo-EM sample preparation, data collection, data processing, model building and refinement. R.M.W.,
S.R. and A.R. performed the data analysis. F.J. and L.H. performed cross-linking mass spectrometry. A.R. and J.H. prepared the figures. J.H. wrote the paper with assistance from B.V., R.M.W.
and S.R., and input from all authors. CORRESPONDING AUTHOR Correspondence to John Hanna. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. PEER REVIEW PEER
REVIEW INFORMATION _Nature Structural & Molecular Biology_ thanks Youdong Mao and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Dimitris
Typas was the primary editor on this article and managed its editorial process and peer review in collaboration with the rest of the editorial team. ADDITIONAL INFORMATION PUBLISHER’S NOTE
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. EXTENDED DATA EXTENDED DATA FIG. 1 CONSERVATION OF MIDLINE STABILIZING
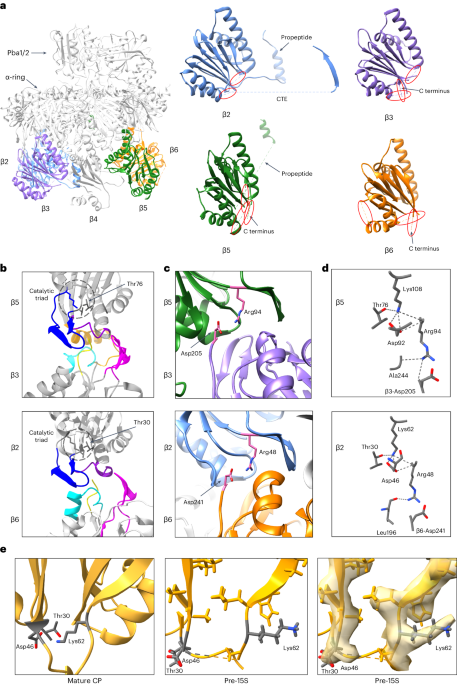
ASPARTATES AND ARGININES. A, Comparison of the C-termini for all 7 yeast β-subunits. Only β3 and β6 end in aspartate. B, Evolutionary conservation of the β3/β5 midline salt bridge residues,
β3-D205 and β5-R94. β5′s nearby catalytic triad residue, D92, is shown for reference. C, Evolutionary conservation of the β2/β6 midline salt bridge residues, β6-D241 and β2-R94. β2′s nearby
catalytic triad residue, D46, is shown for reference. EXTENDED DATA FIG. 2 CRYO-EM CLASSIFICATION OF CP SPECIES. Shown is the processing scheme for classification and refinement of
proteasome species. ‘Junk” classes are colored grey while identifiable species are colored by species. The asterisk denotes cryoDRGN classification - displayed classes show representative
maps generated via k-means clustering and are not necessarily correlated to the particle selection carried out by clustering of the latent embeddings. All other 3D classification steps were
carried out in cryoSPARC. EXTENDED DATA FIG. 3 CRYO-EM DATA ANALYSIS FOR CP SPECIES. A, Representative micrograph of proteasome particles embedded in vitreous ice. Scale bar, 500 Å. A total
of 375,915 individual particles were analyzed. B, Selected 2D class averages of 20S, preholoproteasome, and Blm10-CP particles. C, Proteasome reconstructions were filtered and colored by
local resolution (left panels), gold-standard Fourier shell correlation (FSC) curves from cryoSPARC (center panels), and viewing direction distribution plots (right panels). Resolution was
determined at FSC = 0.143. EXTENDED DATA FIG. 4 COMPARISON OF UMP1 BEFORE AND AFTER CP FUSION. A, Molecular model of Ump1 from pre-15S (PDB: 7LS6) and the β3-D205Δ preholoproteasome. The
first and last resolved residues are shown. B, Overlay of the two models, showing no significant conformational changes. EXTENDED DATA FIG. 5 DETAILED ANALYSIS OF THE Β5-PROPEPTIDE. A,
Comparison of β5 between mature wild-type 20S (PDB: 5CZ4) and the β3-D205Δ preholoproteasome. The mature portions of β5 largely overlap, although there is a slight rotation of β5 towards the
CP midline in the mature 20S, which may reflect final tightening of the CP upon completion of assembly. The first and last resolved residues are indicated. B, Overlay of the two structures
from panel A, highlighting the differences between them. Arrows designate portions of β5 that are unresolved in preholoproteasome. C, Overlay of the molecular model of the β5-propeptide onto
its primary map density, confirming the validity of the model. Boxed panels show close-up views of selected regions and arrows indicate their position in the overall propeptide. EXTENDED
DATA FIG. 6 VALIDATION OF THE Β5 PROPEPTIDE STRUCTURE BY CROSSLINKING MASS SPECTROMETRY. Crosslinking mass spectrometry was performed on β3-D205Δ proteasomes, which identified two crosslinks
involving the β5-propeptide: β5-D61/D62 with β5-D193, and β5-E27 with α5-E131. A third crosslink, β5-K16 with α6-K115, was detected in a prior study13. All three crosslinks are within the
crosslinkable distance for the respective Lys-Lys and Asp/Glu-Asp/Glu crosslinkers, and strongly support the modeled structure of the β5-propeptide. The orange dashed lines indicate the
crosslinked residues. The other colored dashed lines indicate unresolved residues. EXTENDED DATA FIG. 7 GRAPHICAL REPRESENTATION OF THE CROSSLINKING MASS SPECTROMETRY DATA FOR THE
PROPEPTIDES, PBA1/2, AND UMP1. Intersubunit (green lines) and intrasubunit (blue lines) crosslinks are shown for the β-subunit propeptides (panel A), Pba1/2 (panel B), and Ump1 (panel C).
Shaded areas indicate the propeptides. EXTENDED DATA FIG. 8 ANALYSIS OF THE Β1 CATALYTIC TRIAD. Arg38 in β1 shows a similar orientation to the corresponding arginine residues in β5 and β2,
and is hydrogen bonded (dashed lines) to the triad aspartate and lysine residues, suggesting that it may play an important role in β1 activation. However, unlike the situation in β5 and β2,
β1′s Arg38 does not form a salt bridge or even contact its midline partner, β7, suggesting that additional mechanisms may account for β1 activation. SUPPLEMENTARY INFORMATION REPORTING
SUMMARY SOURCE DATA SOURCE DATA FIG. 3G Unprocessed gel and blots. SOURCE DATA FIG. 4B,E Unprocessed gel and blots. RIGHTS AND PERMISSIONS Springer Nature or its licensor (e.g. a society or
other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of
this article is solely governed by the terms of such publishing agreement and applicable law. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Velez, B., Walsh, R.M., Rawson, S.
_et al._ Mechanism of autocatalytic activation during proteasome assembly. _Nat Struct Mol Biol_ 31, 1167–1175 (2024). https://doi.org/10.1038/s41594-024-01262-1 Download citation *
Received: 14 November 2023 * Accepted: 04 March 2024 * Published: 10 April 2024 * Issue Date: August 2024 * DOI: https://doi.org/10.1038/s41594-024-01262-1 SHARE THIS ARTICLE Anyone you
share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the
Springer Nature SharedIt content-sharing initiative