A runx–cbfβ-driven enhancer directs the irf8 dose-dependent lineage choice between dcs and monocytes
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT The transcription factor IRF8 is essential for the development of monocytes and dendritic cells (DCs), whereas it inhibits neutrophilic differentiation. It is unclear how _Irf8_
expression is regulated and how this single transcription factor supports the generation of both monocytes and DCs. Here, we identified a RUNX–CBFβ-driven enhancer 56 kb downstream of the
_Irf8_ transcription start site. Deletion of this enhancer in vivo significantly decreased _Irf8_ expression throughout the myeloid lineage from the progenitor stages, thus resulting in loss
of common DC progenitors and overproduction of Ly6C+ monocytes. We demonstrated that high, low or null expression of IRF8 in hematopoietic progenitor cells promotes differentiation toward
type 1 conventional DCs, Ly6C+ monocytes or neutrophils, respectively, via epigenetic regulation of distinct sets of enhancers in cooperation with other transcription factors. Our results
illustrate the mechanism through which IRF8 controls the lineage choice in a dose-dependent manner within the myeloid cell system. Access through your institution Buy or subscribe This is a
preview of subscription content, access via your institution ACCESS OPTIONS Access through your institution Access Nature and 54 other Nature Portfolio journals Get Nature+, our best-value
online-access subscription $32.99 / 30 days cancel any time Learn more Subscribe to this journal Receive 12 print issues and online access $209.00 per year only $17.42 per issue Learn more
Buy this article * Purchase on SpringerLink * Instant access to full article PDF Buy now Prices may be subject to local taxes which are calculated during checkout ADDITIONAL ACCESS OPTIONS:
* Log in * Learn about institutional subscriptions * Read our FAQs * Contact customer support SIMILAR CONTENT BEING VIEWED BY OTHERS PIVOTAL ROLE OF DPYSL2A IN KLF4-MEDIATED MONOCYTIC
DIFFERENTIATION OF ACUTE MYELOID LEUKEMIA CELLS Article Open access 20 November 2020 THE EPIGENETIC PIONEER EGR2 INITIATES DNA DEMETHYLATION IN DIFFERENTIATING MONOCYTES AT BOTH STABLE AND
TRANSIENT BINDING SITES Article Open access 10 March 2021 THPOK IS A CRITICAL MULTIFACETED REGULATOR OF MYELOID LINEAGE DEVELOPMENT Article 20 July 2023 DATA AVAILABILITY The data supporting
the findings of this study are available from the corresponding authors (T.T. and A.N.) upon reasonable request. The sequencing data generated in this study were deposited in the Gene
Expression Omnibus database (https://www.ncbi.nlm.nih.gov/geo/). RNA-seq data, ChIP-seq data and ATAC-seq data are available at GSE149762. The following data were retrieved from Gene
Expression Omnibus database: RNA-seq for WT pDCs (GSE121446); RNA-seq for WT B cells, WT CD4+ T cells and WT CD8+ T cells (GSE127267); RNA-seq for WT CLPs (GSE109805); RUNX1 ChIP-seq for
Hoxb8-FL cells (GSE84328); RUNX1 ChIP-seq for FDC-P1 cells (GSE81179); RUNX2 ChIP-seq for MA9CL cells (GSE120063); and microarray for IRF8– and IRF8+ LMPPs (GSE113748). The following data
were retrieved from DNA Data Bank of Japan Sequence Read Archive (https://www.ddbj.nig.ac.jp/): H3K27ac ChIP-seq for WT GMP, WT MDP, WT cMoP, WT Ly6C+ monocyte, WT CDP, WT neutrophil,
_Irf8_–/– GMP, _Irf8_–/– MDP and _Irf8_–/– cMoP (PRJDB3411); and IRF8 ChIP-seq for WT MDP (PRJDB3411). The sequencing data and public data used in this study are listed in Supplementary
Table 2. Additionally, the table contains information on the figures associated with these data. Source data are provided with this paper. REFERENCES * Shlyueva, D., Stampfel, G. &
Stark, A. Transcriptional enhancers: from properties to genome-wide predictions. _Nat. Rev. Genet._ 15, 272–286 (2014). Article CAS PubMed Google Scholar * Tamura, T., Kurotaki, D. &
Koizumi, S. Regulation of myelopoiesis by the transcription factor IRF8. _Int. J. Hematol._ 101, 342–351 (2015). Article CAS PubMed Google Scholar * Sichien, D. et al. IRF8
transcription factor controls survival and function of terminally differentiated conventional and plasmacytoid dendritic cells, respectively. _Immunity_ 45, 626–640 (2016). Article CAS
PubMed Google Scholar * Kurotaki, D. et al. IRF8 inhibits C/EBPα activity to restrain mononuclear phagocyte progenitors from differentiating into neutrophils. _Nat. Commun._ 5, 4978
(2014). Article CAS PubMed Google Scholar * Tamura, T., Nagamura-Inoue, T., Shmeltzer, Z., Kuwata, T. & Ozato, K. ICSBP directs bipotential myeloid progenitor cells to differentiate
into mature macrophages. _Immunity_ 13, 155–165 (2000). Article CAS PubMed Google Scholar * Bigley, V. et al. Biallelic interferon regulatory factor 8 mutation: a complex
immunodeficiency syndrome with dendritic cell deficiency, monocytopenia and immune dysregulation. _J. Allergy Clin. Immunol._ 141, 2234–2248 (2018). Article CAS PubMed PubMed Central
Google Scholar * Cytlak, U. et al. Differential IRF8 transcription factor requirement defines two pathways of dendritic cell development in humans. _Immunity_ 53, 353–370 (2020). Article
CAS PubMed PubMed Central Google Scholar * Geissmann, F. et al. Development of monocytes, macrophages, and dendritic cells. _Science_ 327, 656–661 (2010). Article CAS PubMed PubMed
Central Google Scholar * Guilliams, M., Mildner, A. & Yona, S. Developmental and functional heterogeneity of monocytes. _Immunity_ 49, 595–613 (2018). Article CAS PubMed Google
Scholar * Guilliams, M. et al. Dendritic cells, monocytes and macrophages: a unified nomenclature based on ontogeny. _Nat. Rev. Immunol._ 14, 571–578 (2014). Article CAS PubMed PubMed
Central Google Scholar * Dress, R. J. et al. Plasmacytoid dendritic cells develop from Ly6D+ lymphoid progenitors distinct from the myeloid lineage. _Nat. Immunol._ 20, 852–864 (2019).
Article CAS PubMed Google Scholar * Anderson, D. A. III, Dutertre, C.-A., Ginhoux, F. & Murphy, K. M. Genetic models of human and mouse dendritic cell development and function. _Nat.
Rev. Immunol_. https://doi.org/10.1038/s41577-020-00413-x (2020). * Pietras, E. M. et al. Functionally distinct subsets of lineage-biased multipotent progenitors control blood production in
normal and regenerative conditions. _Cell Stem Cell_ 17, 35–46 (2015). Article CAS PubMed PubMed Central Google Scholar * Fogg, D. K. et al. A clonogenic bone marrow progenitor
specific for macrophages and dendritic cells. _Science_ 311, 83–87 (2006). Article CAS PubMed Google Scholar * Yáñez, A. et al. Granulocyte-monocyte progenitors and monocyte-dendritic
cell progenitors independently produce functionally distinct monocytes. _Immunity_ 47, 890–902 (2017). Article PubMed PubMed Central Google Scholar * Weinreb, C., Rodriguez-Fraticelli,
A., Camargo, F. D. & Klein, A. M. Lineage tracing on transcriptional landscapes links state to fate during differentiation. _Science_ 367, eaaw3381 (2020). Article CAS PubMed PubMed
Central Google Scholar * Liu, Z. et al. Fate mapping via Ms4a3-expression history traces monocyte-derived cells. _Cell_ 178, 1509–1525 (2019). Article CAS PubMed Google Scholar *
Hettinger, J. et al. Origin of monocytes and macrophages in a committed progenitor. _Nat. Immunol._ 14, 821–830 (2013). Article CAS PubMed Google Scholar * Kurotaki, D. et al. Essential
role of the IRF8-KLF4 transcription factor cascade in murine monocyte differentiation. _Blood_ 121, 1839–1849 (2013). Article CAS PubMed PubMed Central Google Scholar * Kurotaki, D. et
al. Epigenetic control of early dendritic cell lineage specification by the transcription factor IRF8 in mice. _Blood_ 133, 1803–1813 (2019). Article CAS PubMed PubMed Central Google
Scholar * Kurotaki, D. et al. Transcription factor IRF8 governs enhancer landscape dynamics in mononuclear phagocyte progenitors. _Cell Rep._ 22, 2628–2641 (2018). Article CAS PubMed
Google Scholar * Grajales-Reyes, G. E. et al. Batf3 maintains autoactivation of _Irf8_ for commitment of a CD8α+ conventional DC clonogenic progenitor. _Nat. Immunol._ 16, 708–717 (2015).
Article CAS PubMed PubMed Central Google Scholar * Schonheit, J. et al. PU.1 level-directed chromatin structure remodeling at the _Irf8_ gene drives dendritic cell commitment. _Cell
Rep._ 3, 1617–1628 (2013). Article PubMed Google Scholar * Durai, V. et al. Cryptic activation of an _Irf8_ enhancer governs cDC1 fate specification. _Nat. Immunol._ 20, 1161–1173 (2019).
Article CAS PubMed PubMed Central Google Scholar * Bagadia, P. et al. An _Nfil3_–_Zeb2_–_Id2_ pathway imposes Irf8 enhancer switching during cDC1 development. _Nat. Immunol._ 20,
1174–1185 (2019). Article CAS PubMed PubMed Central Google Scholar * Kawano, Y. et al. Stable lines and clones of long-term proliferating normal, genetically unmodified murine common
lymphoid progenitors. _Blood_ 131, 2026–2035 (2018). Article CAS PubMed Google Scholar * Lara-Astiaso, D. et al. Chromatin state dynamics during blood formation. _Science_ 345, 943–949
(2014). Article CAS PubMed PubMed Central Google Scholar * Chopin, M. et al. Transcription factor PU.1 promotes conventional dendritic cell identity and function via induction of
transcriptional regulator DC-SCRIPT. _Immunity_ 50, 77–90 (2019). Article CAS PubMed Google Scholar * Tamura, T. et al. IFN regulatory factor-4 and -8 govern dendritic cell subset
development and their functional diversity. _J. Immunol._ 174, 2573–2581 (2005). Article CAS PubMed Google Scholar * Lee, J. et al. Lineage specification of human dendritic cells is
marked by IRF8 expression in hematopoietic stem cells and multipotent progenitors. _Nat. Immunol._ 18, 877–888 (2017). Article CAS PubMed PubMed Central Google Scholar * Gonzalez, A.
J., Setty, M. & Leslie, C. S. Early enhancer establishment and regulatory locus complexity shape transcriptional programs in hematopoietic differentiation. _Nat. Genet._ 47, 1249–1259
(2015). Article CAS PubMed PubMed Central Google Scholar * Schlitzer, A. et al. Identification of cDC1- and cDC2-committed DC progenitors reveals early lineage priming at the common DC
progenitor stage in the bone marrow. _Nat. Immunol._ 16, 718–728 (2015). Article CAS PubMed Google Scholar * Factor, D. C. et al. Epigenomic comparison reveals activation of “seed”
enhancers during transition from naive to primed pluripotency. _Cell Stem Cell_ 14, 854–863 (2014). Article CAS PubMed PubMed Central Google Scholar * Kim, S. et al. High amount of
transcription factor IRF8 engages AP1-IRF composite elements in enhancers to direct type 1 conventional dendritic cell identity. _Immunity_ 53, 759–774 (2020). Article CAS PubMed PubMed
Central Google Scholar * Scott, E. W., Simon, M. C., Anastasi, J. & Singh, H. Requirement of transcription factor PU.1 in the development of multiple hematopoietic lineages. _Science_
265, 1573–1577 (1994). Article CAS PubMed Google Scholar * Heinz, S. et al. Simple combinations of lineage-determining transcription factors prime _cis_-regulatory elements required for
macrophage and B cell identities. _Mol. Cell_ 38, 576–589 (2010). Article CAS PubMed PubMed Central Google Scholar * Satpathy, A. T. et al. Runx1 and Cbfβ regulate the development of
Flt3+ dendritic cell progenitors and restrict myeloproliferative disorder. _Blood_ 123, 2968–2977 (2014). Article CAS PubMed PubMed Central Google Scholar * Guo, H., Ma, O., Speck, N.
A. & Friedman, A. D. Runx1 deletion or dominant inhibition reduces _Cebpa_ transcription via conserved promoter and distal enhancer sites to favor monopoiesis over granulopoiesis.
_Blood_ 119, 4408–4418 (2012). Article CAS PubMed PubMed Central Google Scholar * Hamey, F. K. et al. Reconstructing blood stem cell regulatory network models from single-cell molecular
profiles. _Proc. Natl Acad. Sci. USA_ 114, 5822–5829 (2017). Article CAS PubMed PubMed Central Google Scholar * Behrens, K. et al. Runx1 downregulates stem cell and megakaryocytic
transcription programs that support niche interactions. _Blood_ 127, 3369–3381 (2016). Article CAS PubMed Google Scholar * Volk, A. et al. A CHAF1B-dependent molecular switch in
hematopoiesis and leukemia pathogenesis. _Cancer Cell_ 34, 707–723 (2018). Article CAS PubMed PubMed Central Google Scholar * Sigvardsson, M. Transcription factor dose links development
to disease. _Blood_ 120, 3630–3631 (2012). Article CAS PubMed Google Scholar * Wilkinson, A. C., Nakauchi, H. & Gottgens, B. Mammalian transcription factor networks: recent advances
in interrogating biological complexity. _Cell Syst._ 5, 319–331 (2017). Article CAS PubMed PubMed Central Google Scholar * Heinz, S., Romanoski, C. E., Benner, C. & Glass, C. K.
The selection and function of cell type-specific enhancers. _Nat. Rev. Mol. Cell Biol._ 16, 144–154 (2015). Article CAS PubMed PubMed Central Google Scholar * Stadhouders, R., Filion,
G. J. & Graf, T. Transcription factors and 3D genome conformation in cell-fate decisions. _Nature_ 569, 345–354 (2019). Article CAS PubMed Google Scholar * DeKoter, R. P. &
Singh, H. Regulation of B lymphocyte and macrophage development by graded expression of PU.1. _Science_ 288, 1439–1441 (2000). Article CAS PubMed Google Scholar * Rosenbauer, F. et al.
Acute myeloid leukemia induced by graded reduction of a lineage-specific transcription factor, PU.1. _Nat. Genet._ 36, 624–630 (2004). Article CAS PubMed Google Scholar * Tamura, A. et
al. C/EBPβ is required for survival of Ly6C– monocytes. _Blood_ 130, 1809–1818 (2017). Article CAS PubMed PubMed Central Google Scholar * Friedman, A. D. C/EBPα in normal and malignant
myelopoiesis. _Int. J. Hematol._ 101, 330–341 (2015). Article CAS PubMed PubMed Central Google Scholar * Briscoe, J. & Small, S. Morphogen rules: design principles of
gradient-mediated embryo patterning. _Development_ 142, 3996–4009 (2015). Article CAS PubMed PubMed Central Google Scholar * Naito, Y., Hino, K., Bono, H. & Ui-Tei, K. CRISPRdirect:
software for designing CRISPR/Cas guide RNA with reduced off-target sites. _Bioinformatics_ 31, 1120–1123 (2015). Article CAS PubMed Google Scholar * Hashimoto, M. & Takemoto, T.
Electroporation enables the efficient mRNA delivery into the mouse zygotes and facilitates CRISPR/Cas9-based genome editing. _Sci. Rep._ 5, 11315 (2015). Article CAS PubMed PubMed Central
Google Scholar * Sakai, K. et al. _Usp26_ mutation in mice leads to defective spermatogenesis depending on genetic background. _Sci. Rep._ 9, 13757 (2019). Article PubMed PubMed Central
Google Scholar * Tamura, T., Thotakura, P., Tanaka, T. S., Ko, M. S. & Ozato, K. Identification of target genes and a unique _cis_ element regulated by IRF-8 in developing
macrophages. _Blood_ 106, 1938–1947 (2005). Article CAS PubMed PubMed Central Google Scholar * Naldini, L., Blomer, U., Gage, F. H., Trono, D. & Verma, I. M. Efficient transfer,
integration, and sustained long-term expression of the transgene in adult rat brains injected with a lentiviral vector. _Proc. Natl Acad. Sci. USA_ 93, 11382–11388 (1996). Article CAS
PubMed PubMed Central Google Scholar * Ban, T. et al. Lyn kinase suppresses the transcriptional activity of IRF5 in the TLR-MyD88 pathway to restrain the development of autoimmunity.
_Immunity_ 45, 319–332 (2016). Article CAS PubMed Google Scholar * Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. _Bioinformatics_ 29, 15–21 (2013). Article CAS PubMed
Google Scholar * Li, B., Ruotti, V., Stewart, R. M., Thomson, J. A. & Dewey, C. N. RNA-Seq gene expression estimation with read mapping uncertainty. _Bioinformatics_ 26, 493–500 (2010).
Article PubMed Google Scholar * Corces, M. R. et al. Lineage-specific and single-cell chromatin accessibility charts human hematopoiesis and leukemia evolution. _Nat. Genet._ 48,
1193–1203 (2016). Article CAS PubMed PubMed Central Google Scholar * Langmead, B., Trapnell, C., Pop, M. & Salzberg, S. L. Ultrafast and memory-efficient alignment of short DNA
sequences to the human genome. _Genome Biol._ 10, R25 (2009). Article PubMed PubMed Central Google Scholar * Saldanha, A. J. Java Treeview–extensible visualization of microarray data.
_Bioinformatics_ 20, 3246–3248 (2004). Article CAS PubMed Google Scholar * Whyte, W. A. et al. Master transcription factors and mediator establish super-enhancers at key cell identity
genes. _Cell_ 153, 307–319 (2013). Article CAS PubMed PubMed Central Google Scholar * Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting
genome-wide expression profiles. _Proc. Natl Acad. Sci. USA_ 102, 15545–15550 (2005). Article CAS PubMed PubMed Central Google Scholar Download references ACKNOWLEDGEMENTS The authors
thank M. Ichino, I. Harada, M. Yoshinari, S. Honma, H. Sato, G. R. Sato and M. Tachikawa at Yokohama City University for their help with the experiments. This work was supported by
Grants-in-Aid for Scientific Research from the Japan Society for the Promotion of Science/Ministry of Education, Culture, Sports, Science and Technology (MEXT; grant nos. 18K19345 and
15H04860 to T.T. and 19K07372 to A.N.); a Uehara Memorial Foundation Research Grant (to T.T.); a Japanese Society of Hematology Research Grant (to T.T.); and the MEXT Joint Usage/Research
Center Program at the Advanced Medical Research Center, Yokohama City University (funding for Y.S., T.K. and T.T.). AUTHOR INFORMATION Author notes * These authors contributed equally:
Koichi Murakami, Haruka Sasaki, Akira Nishiyama. AUTHORS AND AFFILIATIONS * Department of Immunology, Yokohama City University Graduate School of Medicine, Kanagawa, Japan Koichi Murakami,
Haruka Sasaki, Akira Nishiyama, Daisuke Kurotaki, Wataru Kawase, Tatsuma Ban & Tomohiko Tamura * Advanced Medical Research Center, Yokohama City University, Kanagawa, Japan Koichi
Murakami, Jun Nakabayashi & Tomohiko Tamura * Laboratory of Stem Cell Biology, Department of Biosciences, Kitasato University School of Science, Kanagawa, Japan Satoko Kanzaki, Yoichi
Sekita & Tohru Kimura * Department of Stem Cell and Immune Regulation, Yokohama City University Graduate School of Medicine, Kanagawa, Japan Hideaki Nakajima * Program in Genomics of
Differentiation, Eunice Kennedy Shriver National Institute of Child Health and Human Development, NIH, Bethesda, MD, USA Keiko Ozato Authors * Koichi Murakami View author publications You
can also search for this author inPubMed Google Scholar * Haruka Sasaki View author publications You can also search for this author inPubMed Google Scholar * Akira Nishiyama View author
publications You can also search for this author inPubMed Google Scholar * Daisuke Kurotaki View author publications You can also search for this author inPubMed Google Scholar * Wataru
Kawase View author publications You can also search for this author inPubMed Google Scholar * Tatsuma Ban View author publications You can also search for this author inPubMed Google Scholar
* Jun Nakabayashi View author publications You can also search for this author inPubMed Google Scholar * Satoko Kanzaki View author publications You can also search for this author inPubMed
Google Scholar * Yoichi Sekita View author publications You can also search for this author inPubMed Google Scholar * Hideaki Nakajima View author publications You can also search for this
author inPubMed Google Scholar * Keiko Ozato View author publications You can also search for this author inPubMed Google Scholar * Tohru Kimura View author publications You can also search
for this author inPubMed Google Scholar * Tomohiko Tamura View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS K.M., H.S., A.N. and T.T.
designed the study. K.M., H.S., A.N., D.K., W.K., T.B., S.K. and Y.S. conducted the experiments; K.M., H.S., A.N., J.N. and T.T. analyzed the data; K.M., A.N. and T.T. wrote the manuscript;
K.O. provided key resources; H.N., K.O. and T.K. provided intellectual input; and T.T. supervised the project. K.M., H.S. and A.N. contributed equally to this work. CORRESPONDING AUTHORS
Correspondence to Akira Nishiyama or Tomohiko Tamura. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION PEER REVIEW INFORMATION
_Nature Immunology_ thanks Venetia Bigley, Charlotte Scott, Alberto Yáñez and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports
are available. L. A. Dempsey was the primary editor on this article and managed its editorial process and peer review in collaboration with the rest of the editorial team. PUBLISHER’S NOTE
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. EXTENDED DATA EXTENDED DATA FIG. 1 MODELS OF MYELOID CELL
DIFFERENTIATION IN WT AND _IRF8_−/− MICE. _Irf8_ starts to be expressed at the MPP stage and its expression sharply increases in MDPs. The expression of _Irf8_ further increases as cells
differentiate into the DC lineage, while remaining relatively low or downregulated in the monocytic lineage. Neutrophils do not express _Irf8_. _Irf8_–/– mice lack Ly6C+ monocytes, CDPs,
pDCs, and cDC1s. _Irf8_–/– mononuclear phagocyte progenitors accumulate and aberrantly give rise to neutrophils. MPPs include both MPP3s and MPP4s/LMPPs that express low amounts of _Irf8_.
The dashed lines denote disputed pathways. Cell populations affected by the enhancer or gene deletion are highlighted in green. EXTENDED DATA FIG. 2 FLOW CYTOMETRIC ANALYSIS OF BONE MARROW
AND SPLEEN CELLS. A-H, Representative FACS plots of HSPCs (A), myeloid progenitors (B), CLPs (C), mononuclear phagocyte progenitors (D), and cMoPs (E) in bone marrow and those of monocytes
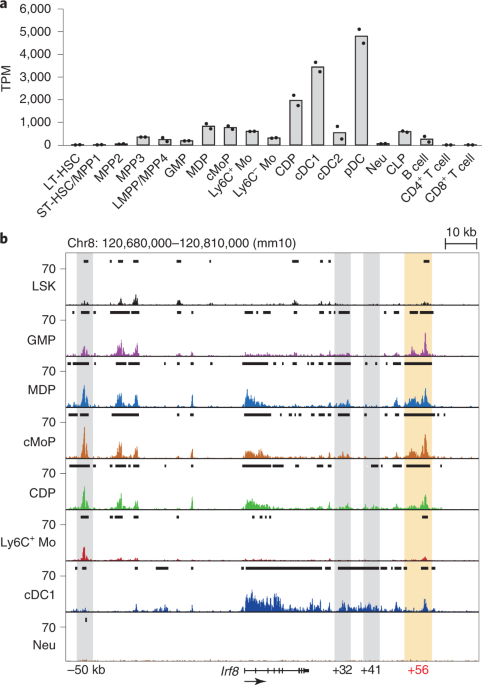
and neutrophils (F), cDCs (G), and pDCs (H) in spleens. EXTENDED DATA FIG. 3 CREATION OF _IRF8_ ENHANCER-NULL MICE BY CRISPR/CAS9 GENOME EDITING. A, The Genome Browser image of the regions
deleted in each enhancer-null mouse strain. B, Representative cropped gel images of genomic PCR confirming the deletion. Primer sets are indicated. Data are representative of over 20
independent experiments for each genotype, which yielded similar results. Full scans are shown in Source Data. C, The Genome Browser image of input DNA data at the _Irf8_ gene locus on ∆+56
cMoP in ChIP-seq analysis. Gray boxes indicate known enhancers at –50, +41, and +32 kb. Source data EXTENDED DATA FIG. 4 THE _IRF8_ +56 KB REGION REGULATES _IRF8_ EXPRESSION AND CELL FATE.
A, Representative FACS plots of pre-cDC1s, pre-cDC2s, pre-DCs, and Ly6C– monocytes analyzed in Fig. 2a. B, IFN-α and IFN-β production by pDCs isolated from WT or ∆+56 mice followed by
overnight stimulation with poly(U) (1.0 µg/mL) or CpG-A (10 µM) (_n_ = 3 mice per genotype). Data are representative of two independent experiments, which yielded similar results. C, In
vitro culture of MDPs. Ten thousand MDPs from WT, ∆+56, and _Irf8_–/– were cultured with Flt3L for 5 days. Representative FACS plots of DC subsets (upper panels) and their absolute cell
numbers (lower panels) produced in the culture are shown (total _n_ = 4 cell cultures). The data were pooled from two independent experiments. D,E, Bone marrow chimera experiments. WT or
∆+56 HSPCs (c-Kit+, 3.0 × 105 cells) were transplanted into irradiated mice (CD45.1+) together with 2.0 × 105 competitor WT whole bone marrow cells (CD45.1+). Cells were analyzed 2 months
after transplantation by FACS and RT-qPCR. In (D), absolute numbers of progenitor populations in bone marrow and differentiated cells in spleens derived from WT or ∆+56 donor cells are
shown. The data were pooled from two independent experiments (total _n_ = 6 mice per genotype). In (E), _Irf8_ mRNA expression in donor-derived bone marrow progenitor populations (total _n_
= 3 mice per genotype). The data were pooled from two independent experiments. Data in B, C (lower panels), D and E are shown as mean + SD. * _P_ < 0.05, ** _P_ < 0.01, *** _P_ <
0.001 (two-tailed Student’s _t_ test) with a fold-change greater than 1.5 or less than 0.66. The exact _P_ values are provided in Source Data. N.D., not detected in (B) and not determined in
(E). Source data EXTENDED DATA FIG. 5 PHENOTYPES OF THE MICE DEVOID OF EITHER THE –50 KB OR +32 KB _IRF8_ ENHANCER. A-D, ∆–50 (A,B) and ∆+32 mouse (C,D) strains were analyzed by FACS and
RT-qPCR. Absolute cell numbers of progenitor populations in bone marrow and differentiated cells in spleens are shown in (A) and (C). The data were pooled from two independent experiments
(total _n_ = 3 mice per genotype for pre-cDC1s, pre-cDC2s, and pre-DCs; total _n_ = 4 mice per genotype for CD43+ Ly6C– monocytes, CD43– Ly6C– cells, and the other cell types of ∆–50 and
∆+32 mice; and total _n_ = 6 mice for the other cell types of WT and _Irf8_–/– mice). _Irf8_ mRNA expression in the indicated cell populations are shown in (B) and (D). The data were pooled
from two independent experiments (total _n_ = 3 mice per genotype except for B cells of ∆–50 mice; _n_ = 2 mice for B cells of ∆–50 mice). All data in Extended Data Figure 5 are shown as
mean + SD. * _P_ < 0.05, ** _P_ < 0.01, *** _P_ < 0.001 (two-tailed Student’s _t_ test) with a fold-change greater than 1.5 or less than 0.66. The exact _P_ values are provided in
Source Data. N.D., not determined. Source data EXTENDED DATA FIG. 6 EXPRESSION OF GFP, EXOGENOUS IRF8, AND ENDOGENOUS IRF8. A, Representative FACS plots of immunostaining for IRF8 in
_Irf8_–/– c-Kit+ cells transduced with a bicistronic retrovirus expressing IRF8 and GFP for two days. Cells in the lower and upper quarters were sorted into GFPlow and GFPhi populations
(left panels). IRF8 expression concentrations in these populations are shown (right panel, _n_ = 3 cell cultures). Data are representative of two independent experiments, which yielded
similar results. B, IRF8 expression in bone marrow progenitor cells and splenic differentiated cells from WT and ∆+56 mice (_n_ = 3 mice per genotype). Data are representative of two
independent experiments, which yielded similar results. ΔMFI was calculated by subtracting the background MFI with isotype control IgG1. Data in A (right panel) and B are presented as mean +
SD; * _P_ < 0.05, ** _P_ < 0.01, *** _P_ < 0.001 (two-tailed Student’s _t_ test). The exact _P_ values are provided in Source Data. N.D., not determined. Source data EXTENDED DATA
FIG. 7 DYNAMICS OF MDP ENHANCERS IN WT, ∆+56, AND _IRF8_−/− MICE. A, Box plots of the normalized H3K27ac ChIP-seq tag densities within the clusters 2, 3, 4, and 5 identified in Fig. 4a in
the indicated cell types from WT, ∆+56, and _Irf8_–/– mice (horizontal lines within the box, median; the lower and upper ends of the box, 25th [Q1] and 75th [Q3] percentiles; the minimum
limit of whiskers, minimum value or Q1 − 1.5× interquartile range [IQR]; the maximum limit of whiskers, maximum value or Q3 + 1.5× IQR). The _P_ values were calculated by the paired
two-tailed _t_ test. The exact _P_ values are provided in Source Data. B, A heat map illustrating normalized enrichment scores (NES) of GSEA for the genes nearest to the regions in the
clusters 2, 3, 4, and 5. Each box shows a GSEA NES that compares the hematopoietic population on its left side with that on its upper side. mRNA expression data were obtained by RNA-seq. A
positive NES means a greater value in the left-hand population. NaN, not-a-number. C, mRNA expression of the representative genes from the clusters 2, 3, 4, and 5 identified in Fig. 4a. The
indicated cell types from WT, ∆+56, and _Irf8_–/– mice were analyzed by RNA-seq (_n_ = 2 biologically independent samples per population). D, Expression of _Klf4_ mRNA analyzed by RNA-seq in
the indicated cell types from WT, ∆+56, and _Irf8_–/– mice (_n_ = 2 biologically independent samples per population). N.D., not determined. Data in C and D are presented as mean. Source
data EXTENDED DATA FIG. 8 EXPRESSION OF GENES ENCODING TFS CO-OPERATING WITH IRF8. mRNA expression amounts of the indicated genes in WT Ly6C+ monocytes and cDC1s determined by RNA-seq (_n_ =
2 biologically independent samples per population). Data are shown as mean. Source data EXTENDED DATA FIG. 9 RUNX–CBFΒ REGULATES THE DEVELOPMENT OF CDCS. A, Genome Browse images of RUNX1
and RUNX2 ChIP-seq tags in HSPC cell lines around the _Irf8_ gene and enhancers, retrieved from previous reports39,40,41. ATAC-seq data on WT MDPs newly obtained in this study (_n_ = 2
biologically independent samples, each using 1 mouse) and H3K27ac ChIP-seq data on WT MDPs retrieved from our previous publication21 (_n_ = 2 biologically independent samples, each using 20
mice) are shown for reference. B,C, WT LSK cells were transduced with a lentivirus encoding shRNA against _Cbfb_ and cultured with SCF and Flt3L as in Fig. 7. Representative FACS plots on
day 7 are shown in (B). The percentages of MDPs and CDPs (day 5, _n_ = 3 cell cultures), cDCs, pDCs, and monocytes/macrophages (day 7, _n_ = 4 cell cultures) among GFP+ cells are shown in
(C). Data are representative of two independent experiments, which yielded similar results. The bar graphs are shown as mean + SD. * _P_ < 0.05, ** _P_ < 0.01, *** _P_ < 0.001
(two-tailed Student’s _t_ test). The exact _P_ values are provided in Source Data. D, Representative histograms of reporter assays in MDPs shown in Fig. 7f. E, mRNA expression of _Runx1_,
_Runx2_, _Runx3_, and _Cbfb_ analyzed by RNA-seq in the indicated cell types from WT mice (_n_ = 2 biologically independent samples). Data are shown as mean. F, Normalized microarray
intensities of _Runx1_, _Runx2_, _Runx3_, _Cbfb_, and _Irf8_ in IRF8– and IRF8+ LMPPs (_n_ = 2 biologically independent samples). Data are shown as mean. G, A Genome Browser image of
sequence conservation at the human _IRF8_ gene locus against the mouse genome. The region corresponding to mouse +56 kb _Irf8_ enhancer (dotted line) and the region used for the +56 kb
enhancer reporter assay (orange) are indicated. Positions of the two RUNX motifs are shown as blue arrow lines. Mac, macrophage; NC, negative control. Source data EXTENDED DATA FIG. 10 THE
PROPOSED MODEL OF THE IRF8-DOSE DEPENDENT MYELOID LINEAGE CHOICE. A, Schematic models of the phenotypes of three mouse strains devoid of the +56, +32 or –50 kb _Irf8_ enhancer. The models
are described as in Extended Data Fig. 1. Phenotypes of WT and _Irf8_–/– mice are also displayed for comparison. B, Proposed model: The RUNX–CBFβ-driven _Irf8_ + 56 kb enhancer induces early
IRF8 expression in myeloid progenitors. The +56 kb enhancer-mediated high IRF8 expression in myeloid progenitor cells is essential for the development of CDPs and cDC1s, whereas low IRF8
expression in myeloid progenitor cells preferentially induces monopoiesis. The absence of IRF8 expression leads to differentiation into neutrophils (Neu). The lineage choice is
epigenetically determined in an IRF8 dose-dependent manner via cooperation or antagonism with other TFs to activate distinct sets of downstream enhancers. TFs, transcription factors.
SUPPLEMENTARY INFORMATION REPORTING SUMMARY PEER REVIEW INFORMATION SUPPLEMENTARY TABLE 1 Oligonucleotides used in this study. SUPPLEMENTARY TABLE 2 Datasets used in this study. SOURCE DATA
SOURCE DATA FIG. 1 Statistical source data with exact _P_ values. SOURCE DATA FIG. 2 Statistical source data with exact _P_ values. SOURCE DATA FIG. 3 Statistical source data with exact _P_
values. SOURCE DATA FIG. 4 Statistical source data with exact _P_ values. SOURCE DATA FIG. 5 Statistical source data with exact _P_ values. SOURCE DATA FIG. 7 Statistical source data with
exact _P_ values. SOURCE DATA EXTENDED DATA FIG. 3 Full scans of gel images for genomic PCR. SOURCE DATA EXTENDED DATA FIG. 4 Statistical source data with exact _P_ values. SOURCE DATA
EXTENDED DATA FIG. 5 Statistical source data with exact _P_ values. SOURCE DATA EXTENDED DATA FIG. 6 Statistical source data with exact _P_ values. SOURCE DATA EXTENDED DATA FIG. 7
Statistical source data with exact _P_ values. SOURCE DATA EXTENDED DATA FIG. 8 Statistical source data with exact _P_ values. SOURCE DATA EXTENDED DATA FIG. 9 Statistical source data with
exact _P_ values. RIGHTS AND PERMISSIONS Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Murakami, K., Sasaki, H., Nishiyama, A. _et al._ A RUNX–CBFβ-driven enhancer directs
the _Irf8_ dose-dependent lineage choice between DCs and monocytes. _Nat Immunol_ 22, 301–311 (2021). https://doi.org/10.1038/s41590-021-00871-y Download citation * Received: 22 May 2020 *
Accepted: 11 January 2021 * Published: 18 February 2021 * Issue Date: March 2021 * DOI: https://doi.org/10.1038/s41590-021-00871-y SHARE THIS ARTICLE Anyone you share the following link with
will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt
content-sharing initiative