Serotonin and dopamine modulate aging in response to food odor and availability
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT An organism’s ability to perceive and respond to changes in its environment is crucial for its health and survival. Here we reveal how the most well-studied longevity intervention,
dietary restriction, acts in-part through a cell non-autonomous signaling pathway that is inhibited by the presence of attractive smells. Using an intestinal reporter for a key gene induced
by dietary restriction but suppressed by attractive smells, we identify three compounds that block food odor effects in _C. elegans_, thereby increasing longevity as dietary restriction
mimetics. These compounds clearly implicate serotonin and dopamine in limiting lifespan in response to food odor. We further identify a chemosensory neuron that likely perceives food odor,
an enteric neuron that signals through the serotonin receptor 5-HT1A/SER-4, and a dopaminergic neuron that signals through the dopamine receptor DRD2/DOP-3. Aspects of this pathway are
conserved in _D. melanogaster_. Thus, blocking food odor signaling through antagonism of serotonin or dopamine receptors is a plausible approach to mimic the benefits of dietary restriction.
SIMILAR CONTENT BEING VIEWED BY OTHERS OLFACTORY PERCEPTION OF FOOD ABUNDANCE REGULATES DIETARY RESTRICTION-MEDIATED LONGEVITY VIA A BRAIN-TO-GUT SIGNAL Article 15 March 2021 DIETARY _E.
COLI_ PROMOTES AGE-DEPENDENT CHEMOTAXIS DECLINE IN _C. ELEGANS_ Article Open access 06 March 2024 A NEUROTRANSMITTER PRODUCED BY GUT BACTERIA MODULATES HOST SENSORY BEHAVIOUR Article 17 June
2020 INTRODUCTION Rapid advances in aging research have identified several conserved signaling pathways that influence aging in organisms across taxa1. Recent work shows that many of these
“longevity pathways” act through cell non-autonomous signaling mechanisms2,3. These pathways utilize sensory cells—frequently neurons—to signal to peripheral tissues and promote survival
during the presence of external stress. Importantly, this neuronal activation of stress response pathways, through either genetic modification or exposure to environmental stress, is often
sufficient to improve health and longevity. Despite mounting evidence that neuronal signaling can influence multiple longevity pathways, less is known about the cells and molecules that
propagate these signals. Biogenic amines are among the most well-studied and conserved neuronal signaling molecules4,5. Specifically, serotonin and dopamine play well-defined roles in
behavior and physiology. However, their role in aging is less well understood. Several recent studies implicate serotonin as an important signal in multiple _C. elegans_ longevity pathways
including the response to heat shock6, hypoxia7 and dietary restriction (DR)8,9. _tph-1_ (WormBase, ZK1290.2), the rate-limiting enzyme for serotonin synthesis, is involved in food abundance
sensing and regulates food deprivation-mediated longevity through both the ADF and NSM neurons8. Perception of food smell dampens the longevity benefit of DR10,11,12. A recent report also
shows that _tph-1_ knockout mutants do not respond to the food smell suppression of DR-mediated longevity, and ADF neuron activity is involved in response to food smell9. Dopaminergic
signaling is associated with physical activity in humans and loss of this signaling decreases lifespan in mice13 and blocks lifespan extension in nematodes14. Serotonin and dopamine levels
both decrease with age across species15,16, consistent with these signaling pathways promoting healthy aging. A pharmacological screen for extending _C. elegans_ lifespan has identified that
compounds modulating serotonin and dopamine signaling promote longevity17. Despite rigorous study and clinical use of drugs that modify serotonin and dopamine signaling, our understanding
of their complex actions and potential interaction is far from complete. Dietary restriction (DR) is the most well-studied and consistent intervention known to improve health and longevity
in organisms ranging from single-celled yeast to primates18. DR leads to improved cell survival and stress resistance, complex intracellular signaling events, metabolic changes, and
increased activity in multiple organisms. Nematode flavin-containing monooxygenase-2 (encoded by _fmo-2_, WormBase, K08C7.5) is necessary and sufficient to increase health and longevity
downstream of DR7. FMOs are highly conserved proteins that are also induced in multiple mammalian models with increased lifespan, including DR19,20. Having previously identified a role for
_fmo-2_ in aging, we wondered whether DR cell non-autonomously regulates _fmo-2_ induction and whether perception of food through biogenic amines could be involved in the subsequent
signaling pathway. In this work, we use _fmo-2_ induction to interrogate the cell non-autonomous DR signaling pathway and how it is affected by food odor. We find that DR induces _fmo-2_
cell non-autonomously to increase longevity through a pathway that involves decreased serotonin signaling from NSM serotonergic neurons and decreased dopamine signaling from dopaminergic
neurons. This process is abrogated by food odor sensed by AWC chemosensory neurons and can be mimicked through small molecules that antagonize biogenic amine signaling. Lastly, we find that
some of these processes are conserved in fruit flies, suggesting that blocking the perception of food is a plausible approach to mimicking DR. RESULTS ATTRACTANT FOOD ODOR REPRESSES _FMO-2_
TO LIMIT LONGEVITY We developed an integrated single-copy _mCherry_ reporter driven by the _fmo-2_ promoter to measure _fmo-2_ induction. The reporter is primarily expressed in the intestine
and responds to stimuli previously reported to induce _fmo-2_, including DR. As an intestinal protein21, we expected that _fmo-2_ would likely be induced cell autonomously by the change in
nutrient intake under DR. To test this hypothesis, we asked whether the perception of food smell by worms in the absence of eating can abrogate the induction of _fmo-2_. Using a “sandwich
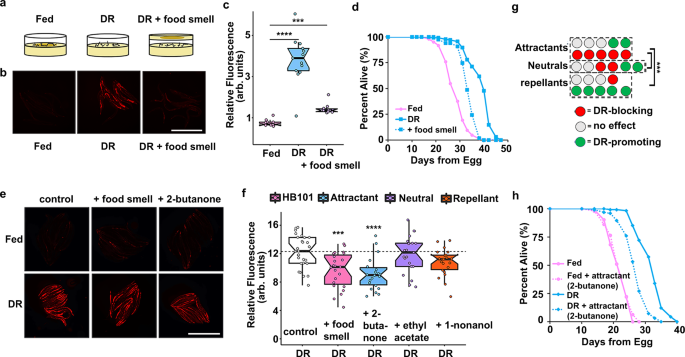
plate” assay as described in Fig. 1a, we were surprised to find a significant reduction in _fmo-2_ induction when worms could smell but not eat food (Fig. 1b, c, _C. elegans_ strains used in
this study are listed in Supplementary Data 1). This reduction is consistent with a model in which increased _fmo-2_ mediates the increase in longevity from DR, as food smell significantly
abrogates lifespan extension by DR, (Fig. 1d, all lifespan replicates in this study are listed in Supplementary Data 2) similar to previous findings9,10,11,12. We also find that live
bacteria are required to abrogate _fmo-2_ induction, as the presence of bacteria killed with 0.5% paraformaldehyde22 does not prevent DR from inducing _fmo-2_ expression (Supplementary Fig.
S1a, b). Since intestinal cells are not known to perceive external environmental cues such as smell, these results suggest that _fmo-2_ expression is suppressed when live food is present
through cell non-autonomous signaling. We next wondered what types of odorants worms sense in this pathway. Bacteria secrete hundreds of volatile compounds that are classified in three
categories based on how they promote chemotaxis: attractants, repellants, and neutral compounds23,24,25. We tested whether exposure to any volatile compound secreted from bacteria is
sufficient to block the lifespan-promoting effects of DR or whether compounds identified as attractants and repellants oppositely regulate _fmo-2_ induction. Using compounds derived from
studies of the _E. coli_ strain HB101 in a range of concentrations (Supplementary Data 3), we find that attractants are more likely to suppress DR-mediated induction of _fmo-2_ (Fig. 1e, f)
whereas some neutral and repellant compounds can induce _fmo-2_ under fed conditions (Supplementary Fig. 1c–h). We also find that many attractive compounds suppress _fmo-2_ expression,
consistent with the hypothesis that this pathway is not acting through a single receptor (Fig. 1g, all results in Supplementary Fig. 2a–z). These results support a model in which perception
of attractive odorants secreted by _E. coli_ abrogates the induction of the pro-longevity gene _fmo-2_. To test whether these odors also affect longevity, we exposed worms to one odorant
from each category and measured lifespans +/− DR. Our results show that the attractant shortened DR lifespan only, recapitulating the effect of food smell on DR mediated lifespan; the
neutral odorant did not affect lifespans of fed or DR worms; while the repellant shortened fed and DR lifespans (Fig. 1h, Supplementary Fig. 1i–j). This is consistent with attractive smells
preventing the lifespan-promoting effects of DR, possibly through a neural response to external stimuli that leads to physiological changes in peripheral tissues. SEROTONIN AND DOPAMINE
ANTAGONISTS INDUCE _FMO-2_ TO MIMIC DR LONGEVITY Biogenic amines can regulate pro-longevity pathways and are involved in behavioral changes in response to food6,7,26,27,28. Some biogenic
amines have previously been reported to regulate DR mediated longevity8,9,17,29. We next asked whether neurotransmitters are involved in the _fmo-_2-mediated food odor pathway. Using a
targeted approach focusing on neurotransmitters and their antagonists, we tested for compounds sufficient to prevent the abrogation of _fmo-2_ induction in the presence of food smell
(Supplementary Fig. 3a–d). The biogenic amine neurotransmitter antagonists mianserin (for serotonin) and thioridazine and trifluoperazine (for dopamine) consistently and significantly
restore _fmo-2_ induction to DR levels in the presence of food smell (Fig. 2a–c, Supplementary Fig. 3e, f). Mianserin is a tetracyclic serotonin antagonist that is thought to competitively
bind to serotonergic G protein-coupled receptors (GPCRs)30 while thioridazine and trifluoperazine’s mechanism of action involves blocking dopamine receptors31. Importantly, while each
compound induces _fmo-2_ to a different extent (Fig. 2d, Supplementary Fig. 3g, i), when combined with DR, no antagonist further induced _fmo-2_, suggesting they act in the same pathway
(Fig. 2e, Supplementary Fig. 3h). Diphenyleneiodonium chloride (DPI), an inhibitor of NADPH oxidase, which we identified in an unpublished screen to robustly induce _fmo-2_ additively with
DR, acts as a positive control, and further induces _fmo-2_ when combined with DR (Fig. 2e). Because thioridazine and trifluoperazine act through similar mechanisms and the effects of
thioridazine were more consistent in our studies, we focused further experiments on dopamine antagonism through thioridazine. Together, these results support antagonism of serotonin or
dopamine as partial mimetics of DR in their induction of _fmo-2_. To validate that the induction of _fmo-2_ through biogenic amine antagonism is beneficial for longevity, we next asked
whether these compounds extend lifespan. We find that both mianserin and thioridazine extend lifespan on agar plates in a dose-dependent manner (Fig. 2f–g). Previous studies report that
mianserin also extends lifespan in liquid culture29, but were not replicated on solid agar plates32. We also confirmed that _fmo-2_ is induced at 25 µM mianserin or 25 µM thioridazine
(Supplementary Fig. 3j–k). Since we identified mianserin and thioridazine through their induction of _fmo-2_, and previously found that _fmo-2_ is necessary for DR-mediated lifespan
extension, we next asked whether _fmo-2_ was necessary for the beneficial longevity effects of mianserin or thioridazine. Our results show that the _fmo-2_ loss of function completely blocks
the lifespan effect of mianserin (Fig. 2h) and thioridazine (Fig. 2i). Importantly, we also see that mianserin treatment combined with DR does not further extend lifespan (Supplementary
Fig. 3l). These results are consistent with these compounds mimicking some aspects of DR-signaling, recapitulating part of the DR lifespan extension effect. Collectively, this supports a
model where DR induces _fmo-2_ because of decreased biogenic amine signaling and establishes neuromodulators as a useful tool to decipher where in the signaling pathway a cell, signal, or
receptor plays a role in DR-mediated longevity. ODOR SENSING AWC NEURONS MODULATE FOOD SMELL RESPONSE Our initial results establish that antagonizing serotonin and dopamine signaling leads
to induction of the longevity promoting _fmo-2_ gene and rescue of the negative effects of food smell. Based on this, we hypothesized that the relative lack of food smell during DR leads to
increased longevity through induction of intestinal _fmo-2_. Using this framework, we next sought to better understand how the sensing of bacteria (or lack thereof) is communicated to
intestinal cells during DR. Our initial results, knocking down _unc-13_, a gene required for both synaptic vesicle33 and dense core vesicle exocytosis34, support short-range
neurotransmitters and/or long-range neuropeptides as necessary for _fmo-2_ induction (Supplementary Fig. 4a, b). In _C. elegans_, perception of the external environment is largely regulated
by a specialized organ known as the amphid. A previous report using a solid-liquid DR approach suggested a pathway originating in the ASI amphid neurons35. DAF-7/TGFβ produced by the ASI
neurons modulates DR longevity8,36,37. We first asked whether these cells are required to modulate _fmo-2_ activity during DR. We find that loss of _daf-3_ (WormBase, F25E2.5) or _daf-7_
(WormBase, B0412.2), each necessary for chemoreceptor signaling in the ASI neurons, did not affect the food odor-mediated reduction in _fmo-2_ expression (Supplementary Fig. 4c, d).
Similarly, proper formation of the amphid (_daf-_6, WormBase, F31F6.5) is also not required (Supplementary Fig. 4e, f). However, it is reported that _daf-6_ mutants can still respond to
volatile odorants23. Three sensory neuron pairs, the AWA/B/C, do not possess the ciliated projections lacking in the _daf-6_ mutant23. To test the necessity of these sensory neurons, we
created individual genetic ablation strains by expressing the pro-apoptotic _caspase-3_ gene (NCBI Gene, 836) under promoters of genes specifically expressed in AWA, AWB, or AWC neurons and
crossed them into our _fmo-2_p::mCherry reporter. We find that loss of AWC neurons prevents _fmo-2_ suppression in the presence of food smell during DR (Fig. 3a, b) while AWA and AWB
knockouts still respond to food odor (Supplementary Fig. 4g–j). This result is consistent with AWC neurons perceiving food odor and with the known role of the AWA/B/C neurons in perception
of external stimuli23,38,39. DR SIGNALING ACTS THROUGH A PAIR OF ENTERIC NEURONS Previous studies report that serotonin regulates DR and mianserin mediated longevity in liquid culture17,29.
To further map this pathway that involves the serotonin antagonist mianserin, we first verified that the biogenic amine serotonin is involved in the DR-mediated longevity pathway. We
subjected animals lacking _tph-1_, the rate-limiting enzyme necessary to produce serotonin, to DR and mianserin. _tph-1_ animals are long-lived compared to wild-type40 and are not further
extended by our DR protocol (Fig. 3c) or mianserin treatment (Supplementary Fig. 5a). These data are supported by the abatement of _fmo-2_ induction on DR (Fig. 3d, e) and mianserin
(Supplementary Fig. 5b, c) when animals are subjected to _tph-1_ (RNAi). As post-mitotic animals, _C. elegans_ have a finite number of neurons with discrete connectivity and functions. Three
neuronal pairs normally express _tph-1_41. The hermaphrodite specific motor neurons (HSN) are located along the ventral tail and regulate egg-laying42 whereas two head neuron pairs, the
amphid neurons with dual sensory endings (ADF) and the neurosecretory motor (NSM) neurons, are involved in modifying behavioral states26,43,44. ADF and NSM neurons are also reported to
regulate food abundance sensing and food deprivation-mediated longevity8,9. To investigate the role of these neuron pairs, we utilized _tph-1_ cell-specific knockouts and found that NSM
(Supplementary Fig. 5e) but not ADF (Supplementary Fig. 5d) neurons are necessary for DR-mediated longevity. Cell-specific rescue of _tph-1_ in a _tph-1_ knockout animal suggests that
_tph-1_ expression in NSM (Fig. 3f), but not ADF (Supplementary Fig. 5f) neurons, is sufficient to promote DR-mediated longevity. Moreover, we find rescuing _tph-1_ expression in NSM neurons
recapitulates lifespan suppression under food smell (Supplementary Fig. 5h) and blunts its effects when knocked out in the NSM neurons (Supplementary Fig. 5j). However, _tph-1_ rescue in
ADF neurons does not rescue lifespan suppression by food smell under DR (Supplementary Fig. 5g) but slightly blunts food smell effects when knocked out in ADF (Supplementary Fig. 5i). These
results suggest that NSM neurons are the primary serotonergic neurons in modulating _fmo-2_ induction and longevity in DR +/− food odor, while ADF neurons may play a smaller role in this
pathway as well. A recent study posits that NSM neurons function similar to enteric neurons with neural projections that directly communicate with the pharynx through a pair of acid-sensing
ion channels (ASICs), DEL-3 (WormBase, F26A3.6) and DEL-7 (WormBase, C46A5.2). Signaling through these channels informs the worm to slow locomotion upon contact with food43. These data led
us to wonder whether the longevity effects of DR also require the ASICs to extend lifespan. We find that _del-7_ mutants look phenotypically wild type in their induction of _fmo-2_ and
lifespan extension, in either DR or DR + food smell (Supplementary Fig. 6a–c). Interestingly, _del-3_ mutant worms show abrogated induction of _fmo-2_ under DR and did not diminish _fmo-2_
induction in response to the smell of food (Fig. 3g–h). These _del-3_ mutant animals still exhibit lifespan extension under DR, despite the decreased induction of _fmo-2_, which is not
abrogated by the smell of food (Fig. 3i). Together, these data support a model whereby the enteric NSM neurons release serotonin in response to food odor and the lack of this release extends
longevity. In addition, the ASIC DEL-3 plays a role in the NSM to both behaviorally43 and physiologically respond to food odor and other food signals. DOPAMINERGIC CEP NEURONS ARE LIKELY
INVOLVED IN ODOR SIGNALING Dopamine synthesis is limited to three neuronal pairs (CEP, ADE, and PDE) in hermaphodites45. All three dopaminergic neurons are required for slow movement when
encountering a bacteria lawn46. To test which dopaminergic neurons respond to food odor, we expressed _cat-2_ (WormBase, B0432.5) in CEP, ADE or PDE neurons in the _cat-2_ KO strain lacking
dopamine production. We find that dopamine synthesis is required for _fmo-2_ induction to be suppressed by food odor under DR (Fig. 3j–k) and that rescuing dopamine production using a
promotor expressed in CEP neurons and some additional cells47 restores the suppression of _fmo-2_ induction by food odor (Fig. 3j–k). However, _cat-2_ rescue in ADE or PDE neurons does not
consistently restore the food odor blunting of _fmo-2_ induction (Supplementary Fig. 6d–g). These results suggest dopamine produced from CEP neurons is most likely necessary for food odor
response. MIANSERIN MIMICS DR IN _FMO-2_-MEDIATED LONGEVITY BY ANTAGONIZING SER-4 Prior reports suggest that serotonin receptor orthologs _ser-1_ (WormBase, F59C12.2) and _ser-4_ (WormBase,
Y22D7AR.13) are necessary for the lifespan benefits of mianserin in _C. elegans_48. We hypothesized that a subset of the serotonin receptor orthologs will also be necessary for mianserin and
DR-mediated _fmo-2_ induction. After two generations of RNAi treatment, _ser-1_ and _ser-4_ were the only two receptors that were necessary for _fmo-2_ induction on mianserin (Fig. 4a,
Supplementary Fig. 7a–c) whereas _ser-4_ knockdown most robustly abrogated DR-mediated _fmo-2_ induction (Supplementary Fig. 7d, e). Further, we find that _ser-4_ RNAi slightly but
significantly increases fed lifespan and prevents DR from extending lifespan (Fig. 4b), supporting the hypothesis that mianserin acts as a DR mimetic by antagonizing serotonin signaling that
occurs during feeding. We confirmed these knockdown results and find that _ser-4_ knockout animals do not respond to the suppression of DR-mediated _fmo-2_ induction by food smell (Fig. 4e,
f). Next, to investigate whether this effect is mediated by neuronal signaling or intestinal expression, we rescued _ser-4_ in _ser-4_ knockout animals with tissue-specific promoters and
found that only neuronal _unc-119p::ser-4_ is sufficient to rescue full induction of _fmo-2_ under DR (Fig. 4c, d). This is consistent with serotonergic signaling within the nervous system,
and not directly to the intestine, regulating the response to food and food smell. Single-cell RNA-seq data show _ser-4_ is broadly expressed throughout the nervous system49. To narrow down
the list of potential neurons acting in our pathway, we rescued _ser-4_ expression in distinct neuronal populations (promoters used in Supplementary Data 1). We find _ser-4_ expressed
exclusively in GABAergic neurons is sufficient to rescue food smell suppression of DR-mediated _fmo-2_ induction (Fig. 4e, f). Importantly, rescuing _ser-4_ expression in neurons producing
biogenic amines (Supplementary Fig. 7f, g) or glutamate (Supplementary Fig. 7h, i) does not change _fmo-2_ induction compared to the _ser-4_ KO. These results suggest one or more GABAergic
neurons known to transcribe _ser-4_ is responding to serotonin release when food odor is present. THIORIDAZINE INDUCES _FMO-2_ AND EXTENDS LIFESPAN THROUGH DOP-3/DRD2 Thioridazine is a
compound that antagonizes dopamine receptor D2 (DRD2) in mammals50,51,52, and mimics DR by inducing _fmo-2_ to increase longevity in nematodes (Fig. 2). Based on its role in mammals, we
tested whether nematode DRD2 is involved in DR and mianserin-related _fmo-2_ induction and longevity. When the DRD2 ortholog _dop-3_ (WormBase, T14E8.3) is knocked down by RNAi, _fmo-2_
induction is not affected in fed conditions but its induction by DR is diminished, while its induction by thioridazine is completely abrogated (Fig. 4g, Supplementary Fig. 8a). This result
is consistent with _dop-3_ being required for dopaminergic induction of _fmo-2_. To demonstrate the epistasis of _dop-3_ and _ser-4_ in the signaling pathway, we combined _ser-4_ RNAi with
mianserin and thioridazine treatment. The results show that _ser-4_ depletion blocks _fmo-2_ induction by thioridazine as well as suppresses _fmo-2_ induction by mianserin, as expected (Fig.
4a). Similarly, depletion of _dop-3_ blocks both mianserin and thioridazine from inducing _fmo-2_ (Fig. 4g; Supplementary Fig. 8a)_. dop-3_ knockouts show similar results to _dop-3_ RNAi
depletion (Supplementary Fig. 8a, b). These results support a model where both serotonin and dopamine signaling are epistatic to each other and are each required for full induction of
_fmo-2_ under DR. To test whether DOP-3_/_DRD2 is necessary for lifespan extension by DR and mianserin, we depleted _dop-3_ with RNAi under DR and found that _dop-3_ depletion increases
lifespan but is not further extended by DR (Fig. 4h). Similar to _ser-4_ (Fig. 4e–f), we also confirmed that _dop-3_ knockout worms do not respond to the suppression of _fmo-2_ induction by
food odor under DR (Supplementary Fig. 8c, d). Two dopamine receptor D1 (DRD1) orthologs, _dop-1_ (WormBase, F15A8.5) and _dop-4_ (WormBase, C52B11.3), may also play a role in food odor
response (Supplementary Fig. 8c, d). Since DRD1 is not a primary target of thioridazine50,51,52, and DRD1 (a Gαs-coupled D1-like receptor inducing cAMP53,54) has antagonistic effects of DRD2
(a Gαi/o-coupled D2-like receptor inhibiting cAMP53,54) on behaviors in mammals55 and _C. elegans_56, _dop-1_ and _dop-4_ might be acting in a parallel pathway in food perception. Together,
these results suggest that dopamine and serotonin signaling interactively suppress _fmo-2_ expression to limit lifespan when food and/or odor are present. To further test the target
specificity of mianserin and thioridazine in _C. elegans_, we measured the induction of _fmo-2_ by mianserin or thioridazine in _ser-4_ or _dop-3_ knockout worms (Supplementary Fig. 8E, F).
The results show that _fmo-2_ induction by mianserin or thioridazine is decreased in _ser-4_ or _dop-3_ knockout worms. This further suggests that induction of _fmo-2_ by mianserin and
thioridazine requires both _ser-4_ and _dop-3_. In addition to _ser-4_, the octopamine receptor _ser-3_ (WormBase, K02F2.6) is also antagonized by mianserin and is required for lifespan
extension by mianserin29. Thus we asked whether _ser-3_ is involved in this pathway by testing whether mianserin or thioridazine induces _fmo-2_ through _ser-3_. The results show that
_fmo-2_ induction by mianserin or thioridazine is decreased but not blocked in _ser-3_ knockouts compared to wild type worms (Supplementary Fig. 8e, f). This is consistent with _ser-3_
playing a role in _fmo-2_ induction by mianserin and thioridazine. We interpret this as _ser-3_ acting downstream of both drugs but not as the primary/only downstream receptor. This result
agrees with previous reports that suggest octopamine signaling is downstream of dopamine in the food availability response57,58 and interacts with serotonin in regulating body fat59 and
aversive behaviors60. Based on this, we tested whether octopamine signaling is involved in sensing food odor. We found that the octopamine synthesizing enzyme _tbh-1_ (WormBase, H13N06.6)
and to a lesser extent _ser-3_ knockout animals lack suppression of DR-mediated _fmo-2_ induction in the presence of food odor. This suggests that octopamine is involved in this pathway but
plausibly through multiple octopamine receptors (Supplementary Fig. 8g, h). MIANSERIN EXTENDS _D. MELANOGASTER_ LIFESPAN Having identified serotonin and dopamine antagonism upstream of
_fmo-2_ induction under DR, we were curious whether these relationships might be conserved. Similar to data in worms, recent data in the vinegar fly _D. melanogaster_ show that altered
serotonin signaling can change the ability to assess caloric quality and modulate lifespan27. As we found a narrow range of effective doses in worms (Fig. 2f), we tested a higher dose of
mianserin in vinegar flies (2 mM) for its effect on Fmo2 induction. The resulting data show that both mianserin and fasting (DR) increase expression of fly _fmo-2_ (NCBI Gene 35561)
expression (Fig. 5a), but not _fmo-1_ (NCBI Gene 37814) (Supplementary Fig. 9a). We then asked whether mianserin could also extend lifespan in flies. Using several concentrations, we find a
positive correlation between mianserin dosage and increased lifespan until reaching a detrimental level of serotonin antagonism (Fig. 5b, Supplementary Fig. 9b–d). We also find a comparable
dose response among male and female flies. We note that mianserin treatment does not significantly alter food consumption (Supplementary Fig. 9e, f), as measured by the Fly Liquid-Food
Interaction Counter (FLIC) assay61. Together, these results are consistent with conserved induction of _fmos_ by mianserin and DR, in addition to conserved lifespan extension. DISCUSSION Our
experimental data in _C. elegans_ support a model where dietary restriction (sDR) leads to decreased serotonin release from the enteric NSM neurons and decreased dopamine release, likely
from the CEP neurons. These decreases lead to reduced serotonin binding to the SER-4/5-HT1A receptor and a reduction in dopamine signaling to downstream DOP-3/DRD2 receptors. Subsequently,
the loss of binding to SER-4 and DOP-3 causes a downstream induction of _fmo-2_ and extension of lifespan. It is notable that both SER-4 and DOP-3 receptors are known to dampen adenylyl
cyclase activity when bound, thus the lack of signal will increase the probability of excitement of the cell expressing these receptors. In our working model, we hypothesize that the
presence of food odor acts as a signal sensed by AWC neurons, which signal upstream, downstream, or in parallel with NSM and CEP neurons that release serotonin and dopamine, respectively
(Fig. 5c). The released serotonin and dopamine bind to the serotonin receptor SER-4 on one or more GABAergic neurons and the dopamine receptor DOP-3 respectively to suppress _fmo-2_
induction and longevity (Fig. 5c). We note that the GABAergic neuron, RIS, expresses more _ser-4_ than any other neuron and thus is a good candidate for a role in this signaling pathway62.
In contrast, under sDR, food signals are likely decreased to a level insufficient to excite the neural circuits involving AWC, NSM, and CEP neurons. Decreased serotonin and dopamine release
under DR can be mimicked by serotonin and dopamine antagonists that induce _fmo-2_ and promote longevity (Fig. 5d). Critically, these data highlight that understanding how the nervous system
evaluates and appropriately integrates large amounts of external stimuli, like the availability of food, allows us to target the sensory-integration processes to mimic key aspects of
pro-longevity pathways. A previous report finds that mianserin antagonizes octopamine and serotonin to increase lifespan and that its mechanism overlaps with DR29. Our results reinforce this
previous finding and further identify key neurons, neurotransmitters and receptors that respond to food availability. We also demonstrate FMO-2, a monooxygenase that is sufficient to extend
lifespan in worms7, as a converging downstream effector of serotonin and dopamine signaling for longevity benefits. Our findings suggest that NSM is important for both DR and food odor
effects on lifespans and ADF may play a smaller role as well (Fig. 3f; Supplementary Fig. 5d–j). It is notable that a recent report shows that ADF neuron activity is responsible for food
smell effects on longevity9. This discrepancy may be due to the difference in methodology. We tested the role of serotonin produced by NSM or ADF by both adding the serotonin synthetase
TPH-1 (_tph-1_ cell-specific rescue) and subtracting TPH-1 (_tph-1_ cell-specific knockout) in these neurons to specifically target serotonin production in NSM or ADF neurons. In comparison,
the other study ablated the output of serotonergic neurons using tetanus toxin. Tetanus toxin is a broad signaling inhibitor that will likely affect all signals from serotonergic neurons.
It is also interesting to note that both the NSM neurons and the NSM enriched acid-sensing ion channels (ASICs) DEL-3 are involved in both sensing food odor for longevity (Fig. 3) and food
ingestion in behavioral changes as previously reported43. DEL-3 and DEL-7 are required for NSM activation of post-food encounter slowing, however may not be involved in the behavioral
changes from food odor43. It is intriguing that dopamine and serotonin signaling interactively induce _fmo-2_ and extend lifespan in a common dietary restriction-mediated pathway. In
nematodes, slowing locomotion in the presence of food is thought to be distinctly regulated by mechanosensation leading to dopamine release while dwelling behavior is potentiated by
serotonin46. Significant scientific effort has identified much of the circuitry these neurotransmitters use to promote changes in chemotaxis and egg-laying26,43,63,64,65. Those data suggest
worms can interpret and implement a diverse set of responses to their changing environment. In mammals, SER-4/5-HT1A receptor activation increases dopamine release throughout the brain66,67.
Similarly, recent work shows release of serotonin and dopamine in the human brain influence non-reward-based aspects of cognition and behaviors like decision making68. These findings
support a conserved link between these two neurotransmitters in regulating complex phenotypes like aging. Interestingly, dopamine may have a more complex role in aging. Knockout of dopamine
reuptake receptor _dat-1_, which leads to increased dopamine signaling, and knockout of _cat-2_ that decreases dopamine synthesis, both can shorten lifespans in worms69. Dopamine antagonists
are found to extend worms lifespan in a pharmacological screen17. Our data also show that dopamine antagonists can promote longevity by inducing _fmo-2_ (Fig. 2). It is possible that
dopamine has both positive and negative effects on lifespan depending on the context. We note that although antagonists of dopamine and serotonin signaling induce _fmo-2_ in worms,
decreasing this signaling by _tph-1_, _ser-4_, _dop-3_ or _del-3_ knock down or even AWC ablation is not sufficient to induce _fmo-2_ under fed conditions (Figs. 3d, e, g, h; 4a, g,
Supplementary Fig. 10). While not directly tested here, we hypothesize that two potentially overlapping possibilities could explain this: 1) Because there are multiple sensory inputs for
food perception, loss of just the food smell pathway is not sufficient to produce a meaningful output, and 2) when cells or genes are absent or highly knocked down throughout development,
the signaling networks they regulate undergo rewiring to respond to cues through different pathways. These hypotheses could both play some role in our observed results, and we plan future
research to test them. It is also intriguing that one of these drugs, mianserin, induces Fmo genes in flies. This leads to increased lifespan. Since mianserin treatment extends fly lifespan,
we suspect it acts through a similar mechanism, serotonin antagonism, to mimic DR. This hypothesis is bolstered by _fmo-2_ induction under acute mianserin exposure and fasting, analogous to
what we see in _C. elegans_. We note that in combination with DR, mianserin does not increase longevity of worms any further. It is not known whether FMOs or 5-HT1A receptors are necessary
for mianserin or DR-mediated longevity in flies, but 5-HT2A receptors are necessary for proper food valuation27, suggesting that altering serotonin signaling may prove fruitful in future
studies. Mammals and _C. elegans_ share a single common ancestral Fmo21 and mammalian Fmos share similar homology to _C. elegans fmo-2_, with Fmo5 (NCBI Gene 14263) having the highest %
identity. It will be interesting to investigate whether mianserin is beneficial for health and longevity in mammals. To achieve this goal, it is imperative that we understand the causative
changes of pro-longevity drugs, such as atypical serotonin antagonists, that are known to have pleiotropic effects in humans. In addition to providing the potential for long-term health
benefits, this knowledge will benefit our understanding of serotonin and dopamine signaling networks that affect many human processes and diseases outside of aging. METHODS STRAINS AND
GROWTH CONDITIONS Standard procedures for _C. elegans_ strain maintenance70 were used. In detail, strains were kept at 20 °C in a temperature-controlled incubator and were fed on Escherichia
coli (OP50) seeded on solid nematode growth medium (NGM). Worms were picked or transferred gently by a platinum wire. Additionally, worms were exposed to the smell of OP50 or HB101 when
indicated in the experiments. Supplementary Data 1 includes a list of the strains and RNAi conditions used in this study. All genotypes were confirmed using PCR. _FMO-2P::MCHERRY_ CONSTRUCT
We PCR amplified _mCherry_ from pHG8 and the _fmo-2_ promoter from the worm gDNA under the _fmo-2_ promoter and cloned them into pdonr221 and P4-P1r, respectively. From here, they were
combined using Gateway LR cloning (Invitrogen) to create _fmo-2p_::_mCherry_::unc-54 3’UTR on PCFJ150. AWA/B/C ABLATION CONSTRUCTS We purchased donor plasmid P_mec-18_::caspase-3 (p12)::nz
[TU#813] from Addgene (Plasmid #16082) and P_mec-18_ cz::caspase-3 (p17) [TU#814] from Addgene (Plasmid #16083), and used Gibson cloning (NEB) to replace P_mec-18_ with P_odr-7_. Three
constructs P_odr-7_::caspase-3(p12)::nz, P_odr-7_::cz::caspase-3(p17), and P_odr-7_::GFP were co-injected with fluorescent co-injection marker _myo-3p_::GFP (20 ng/µL) into the
_fmo-2_p::_mCherry_ transcriptional reporter strain to generate AWA genetic ablation strains. Similarly, P_str-1_ was constructed into TU#813 and TU#814 to replace P_mec-18_. Three
constructs P_str-1_::caspase-3(p12)::nz, P_str-1_::cz::caspase-3(p17), and P_str-1_::GFP were co-injected with fluorescent co-injection marker _myo-3p_::GFP (20 ng/µL) into the
_fmo-2_p::_mCherry_ transcriptional reporter strain to generate AWB genetic ablation strains. The AWC genetic ablation strain (oyIs85 [_ceh-36_p::TU#813 + _ceh-36_p::TU#814 + _srtx-1_p::GFP
+ _unc-122_p::DsRed]) is from CGC, PY7502. AWC genetic ablation strain was crossed with _fmo-2_p::_mCherry_. All plasmids were verified via restriction digest and sanger sequencing. ApE
files available upon request. SER-4 AND CAT-2 RESCUE CONSTRUCTS We purchased donor plasmid pPD117.01 from Addgene and used Gibson cloning (NEB) to swap out promoters driving cDNA of
SER-4::SL2::GFP (on backbone) expression. We used the _unc-119_ promoter to target all neurons and the _vha-6_ promoter to target the intestine. We used _unc-47_, _vglu-2_, and _cat-1_
promoters to target SER-4 expression in GABAergic, glutamatergic, and biogenic amine neurons, respectively. Similarly, we constructed promoters driving cDNA of CAT-2::SL2::GFP expression. We
used _swip-10_, _trpa-1_ and _tax-2_ promoters to target CAT-2 expression in CEP, ADE, and PDE neurons, respectively. We used _cat-1_ promoter to rescue CAT-2 expression in all three
dopaminergic neurons (CEP, ADE, and PDE). All plasmids were verified via restriction digest and sanger sequencing. ApE files available upon request. MICROINJECTION Single-copy integration of
_fmo-2p_::_mCherry_::unc-54 3’UTR on PCFJ150 using the ttTi5605 (EG6699) Mos allele was performed as previously described71. Overexpression transgenic animals were generated by injecting
PureLink (Invitrogen) miniprepped DNA clones (~50 ng/µL) with fluorescent co-injection marker _myo-2p_::mNeonGreen (15 ng/µL) or _sur-5p_::sur-5::NLSGFP (20 ng/µL) and junk DNA (up to 100
ng/µL) into gonads of day 1 gravid adult hermaphrodites. Standard protocols were followed to isolate and obtain stable over-expression mutants72. Because transgene expression can vary
substantially, we typically characterized 2–4 independent transgenic lines per experiment. LIFESPAN MEASUREMENTS Lifespans were carried out as previously described with minor
modifications73. Briefly, 20–30 gravid adult animals were placed on NGM plates for a timed egg-lay. After 12–16 h, these animals were removed. Once their progeny reached late L4/early adult
stage, animals were transferred to plates with 33 µL of 150 mM fluorodeoxyuridine (FUdR) and 100 µL of 50 mg/mL Ampicillin per 100 mL NGM to prevent the development of progeny and growth of
bacteria. Roughly 75 worms were placed on each NGM + FUdR plate seeded with concentrated bacteria (×5). A minimum of two plates per strain per condition were used per replicate. Lifespan
plates were transferred periodically during early adulthood to prevent starvation and avoid contamination. Animals were scored as dead and removed from the experiment when they did not move
in response to prodding under a dissection microscope. It is notable that FUdR can extend lifespan and is a confounding factor contributing to lifespan extension74, however, to avoid
matricide under DR, we include FUdR in the lifespan plates in this study. This factor should be considered when interpreting the results. RNAI KNOCKDOWN The RNAi feeding bacteria were
obtained from the Ahringer _C. elegans_ RNAi feeding library. All RNAi plasmids were sequenced to verify the correct target sequence. Animals were exposed to RNAi plates from egg on plates
consisting of NGM supplemented with 1 mM β-D-isothiogalactopyranoside (IPTG) and 25 μg/ml carbenicillin. At late L4 stage of development the animals were transferred to plates containing
freshly seeded RNAi bacteria plus FUdR. RNAi efficiency of genes expressed in neurons is not validated by qPCR but RNAi phenotypes are recapitulated by gene knockouts. It is notable that
RNAi feeding bacteria are HT115 and are reported to modify lifespan compared to OP5075,76. This should be taken into consideration when comparing the lifespan data of RNAi feeding for gene
knock down with the lifespan data of gene mutants fed on OP50. PFA TREATMENT In order to metabolically kill OP50 in food smell assays, bacteria cultures were treated with 0.5% PFA. After 16
h of shaking, 50 mL of the bacteria were aliquoted into 250 mL Erlenmeyer flasks. 32% PFA was added to the flasks to get the desired final PFA concentration (e.g., 781 µL of PFA was added to
get the final concentration of 0.5% PFA). PFA-treated bacteria were shaken at 37 °C for 1 h and then transferred to 50 mL conical tubes, centrifuged and washed with LB five times to remove
residual PFA before seeding. DRUG TREATMENTS Recent reports show improved health outcomes and longevity in nematodes treated with mianserin48, but only in liquid culture32. As our studies
are on agar plates, we modified previous protocols by adding mianserin, thioridazine or trifluoperazine before pouring NGM agar plates. Without proper dosing, these neurotransmitter
antagonists can cause off-target effects like fleeing, especially when combined with DR. All subsequent _C. elegans_ experiments were performed at 50 µM of mianserin and 25 µM of
thioridazine unless otherwise noted. All drugs were purchased from Sigma-Aldrich and were initially dissolved in milliQ water at 2 mM (mianserin) or 100 mM concentration (DRD2 antagonists),
aliquoted, and stored at −20 °C. DIETARY RESTRICTION (DR) LIFESPAN TREATMENTS Lifespan DR assays were performed like other lifespans until day two of adulthood when the worms were
transferred to plates with 109 cfu/ml seeded lawns and transferred every other day four times. This form of DR is termed solid DR (sDR)77. For short-term DR assays, worms were starved for 8
(real-time PCR) or 20 h (slide microscopy). We added 100 µL of 10 mM palmitic acid (Sigma-Aldrich) dissolved in 100% EtOH to the outer rim of the plate to prevent fleeing. ATTRACTANT,
REPELLANT, AND NEUTRAL SMELL TREATMENTS Fed and DR plates were prepared using NGM plates with palmitic acid. Odorants were chosen from previously published work isolating secreted compounds
from the _E. coli_ strain HB10123,24. All concentrations of attractant, repellant, and neutral chemicals were dissolved in 100% ethanol (more details in Supplementary Data 3). A small pad of
NGM agar (2 mL) was poured on the lid of each plate and allowed to solidify before 100 µL of each smell concentration was added to the agar pads. Plates were prepared the day prior to use
to allow the ethanol solutions to dry. Young adult _fmo-2p::mCherry_ worms were placed on fed and DR plates and exposed to each smell for 20 h before fluorescent microscopy images were
taken. SLIDE MICROSCOPY All images in this study were acquired using Leica Application Suite X software and Leica scope with >15 worms/treatment at x6.3 magnification. Worms were
paralyzed in 0.5 M sodium azide (NaN3). Fluorescence mean comparisons were quantified in ImageJ bundled with 64-bit Java 1.8.0 using the polygon tool and saved as macros. Data were plotted
by R version 4.1.0, Microsoft Excel 365, Adobe Photoshop 2021, and Adobe Illustrator 2021. REAL-TIME PCR 500 N2 worms per biological replicate were transferred at young adulthood, 2.5 days
post-hatch, to FuDR plates either seeded with OP50 of fed or DR condition or poured with the addition of 50 µM mianserin or thioridazine. Alternatively, N2 worms were exposed to RNAi plates
from egg on plates consisting of NGM supplemented with 1 mM β-D-isothiogalactopyranoside (IPTG) and 25 μg/ml carbenicillin for two generations. Worms were harvested in 50 µL of M9 and flash
frozen in liquid nitrogen after 8 hours of exposure. Samples were freeze-thawed three times in Trizol reagent (Invitrogen) and RNA was extracted following standard phenol-chloroform
protocols from the manufacturer. Superscript reverse transcriptase II (Invitrogen) was used to synthesize cDNA. 600 ng of cDNA/sample were used with PowerUp SYBR Green Master Mix (Applied
Biosystems) was used in the quantitation with fmo-2 forward primer ACGAAACGAATGAGTCGTCAGT and reverse primer AGAGCAGACAAGAACGCCAT. Canton-S flies were mated and reared on standard food for 2
weeks before separating the flies by sex onto SY10 food with 20 flies/vial. Flies were acclimated to the vials for 24 h before being transferred to SY10 vials coated with 2 mM mianserin or
water (control) or vials containing 2% agar to mimic dietary restriction. After 8 h on these treatments, flies were frozen at −80 °C overnight. Fly heads and bodies were then separated by
vortexing and dissection by forceps (all samples and materials were kept on dry ice throughout). Each treatment contained 3 biological replicates composed of 10 bodies each. Trizol Reagent
(Invitrogen) was used in the RNA extraction, the MultiScribe Reverse Transcriptase kit (Applied Biosystems) was used to synthesize the cDNA, and the real-time PCR analysis used PowerUp SYBR
Green Master Mix (Applied Biosystems) and a StepOne Plus Real-time PCR system (Applied Biosystems) primers of _fmo-1_ (forward primer GCGATAGGATGGGCAAACTG and reverse primer
CCCGGAAGTGGAGCAAATTC) and _fmo-2_ (forward primer CGCAACCAGAAGAAAGCACA and reverse primer TGCTCCTGTACGTGTCCAAT). FLY HUSBANDRY The laboratory stock Canton-S was used in the lifespan and
molecular experiments. Flies were maintained on standard food and housed at 25 °C and 60% relative humidity in a 12:12 h light-dark cycle. FLY SURVIVAL ASSAYS For lifespan measurements,
flies were reared under controlled larval density and collected onto standard food within 24 h of eclosion. Flies were mated for 2–3 days then sorted by sex under light CO2 onto vials
containing standard food used in lifespan experiments (10% sucrose/10% yeast, or SY10), according to well-establish lifespan protocols78. Flies were transferred to fresh food every 2–3 days.
At the beginning of the lifespan, mianserin was dissolved in water at a 1 mM stock concentration and stored at −20 °C. Weekly aliquots were prepared and diluted with water to yield the
final concentrations of 20–80 µM. 100 µL of the drug solution (or water for the control) was added to the top of each vial and kept at room temperature to dry for approximately 2 h before
transferring the flies. STATISTICAL ANALYSES All box plots show individual data points while the box represents SEM (centered on the mean), and whiskers represent 10%/90%. Comparisons
between more than two groups were done using ANOVA. For multiple comparisons, Welch’s Two Sample _t_-test (two-sided) was used, and p values are *_p_ < 0.05, **_p_ < 0.01, ***_p_ <
0.001, and ****_p_ < 0.0001. For lifespan assays, the statistical groupwise and pairwise comparisons among survivorship curves were performed by Online application for survival analysis
(OASIS 2)79. _P_ values were obtained using the log-rank analysis (select pairwise comparisons and group comparisons or interaction studies) as noted. Summary lifespan data, sample size
(_n_), and statistics are included in Supplementary Data 2. Measurements were taken from distinct samples. The box plots display the median by the middle line of the box. The upper boundary
of the box indicates the 75% interquartile range, and the lower boundary indicates the 25% interquartile range. DATA AVAILABILITY All data generated or analyzed during this study are
included in this published article (and its Supplementary Information files). Source data are provided with this paper. REFERENCES * López-Otín, C., Blasco, M. A., Partridge, L., Serrano, M.
& Kroemer, G. The hallmarks of aging. _Cell_ 153, 1194–1217 (2013). Article CAS PubMed PubMed Central Google Scholar * Medkour, Y., Svistkova, V. & Titorenko, V. I.
Cell-Nonautonomous Mechanisms Underlying Cellular and Organismal Aging. _Int Rev. Cell Mol. Biol._ 321, 259–297 (2016). Article CAS PubMed Google Scholar * Miller H. A., Dean E. S.,
Pletcher, S. D. & Leiser S. F. Cell non-autonomous regulation of health and longevity. _Elife_. Dec 2020;9 https://doi.org/10.7554/eLife.62659 * Berger, M., Gray, J. A. & Roth, B. L.
The expanded biology of serotonin. _Annu Rev. Med_ 60, 355–366 (2009). Article CAS PubMed PubMed Central Google Scholar * Arias-Carrión, O., Stamelou, M., Murillo-Rodríguez, E.,
Menéndez-González, M. & Pöppel, E. Dopaminergic reward system: a short integrative review. _Int Arch. Med_ 3, 24 (2010). Article PubMed PubMed Central Google Scholar * Tatum, M. C.
et al. Neuronal serotonin release triggers the heat shock response in C. elegans in the absence of temperature increase. _Curr. Biol._ 25, 163–174 (2015). Article CAS PubMed Google
Scholar * Leiser, S. F. et al. Cell nonautonomous activation of flavin-containing monooxygenase promotes longevity and health span. _Science_. 2015; https://doi.org/10.1126/science.aac9257
* Entchev, E. V. et al. A gene-expression-based neural code for food abundance that modulates lifespan. _Elife_ 4, e06259 (2015). Article CAS PubMed PubMed Central Google Scholar *
Zhang, B., Jun, H., Wu, J., Liu, J. & Xu, X. Z. S. Olfactory perception of food abundance regulates dietary restriction-mediated longevity via a brain-to-gut signal. _Nat. Aging._ 1,
255–268 (2021). Article PubMed PubMed Central Google Scholar * Libert, S. et al. Regulation of Drosophila life span by olfaction and food-derived odors. _Science_ 315, 1133–1137 (2007).
Article CAS ADS PubMed Google Scholar * Smith, E. D. et al. Age- and calorie-independent life span extension from dietary restriction by bacterial deprivation in Caenorhabditis elegans.
_BMC Dev. Biol._ 8, 49 (2008). Article PubMed PubMed Central Google Scholar * Kaeberlein, T. L. et al. Lifespan extension in Caenorhabditis elegans by complete removal of food. _Aging
Cell_ 5, 487–494 (2006). Article CAS PubMed Google Scholar * Grady, D. L. et al. DRD4 genotype predicts longevity in mouse and human. _J. Neurosci._ 33, 286–291 (2013). Article CAS
PubMed PubMed Central Google Scholar * Saharia, K., Kumar, R., Gupta, K., Mishra, S. & Subramaniam, J. R. Reserpine requires the D2-type receptor, dop-3, and the exoribonuclease,
eri-1, to extend the lifespan in C. elegans. _J. Biosci._ 41, 689–695 (2016). Article CAS PubMed Google Scholar * Yin, J. A., Liu, X. J., Yuan, J., Jiang, J. & Cai, S. Q. Longevity
manipulations differentially affect serotonin/dopamine level and behavioral deterioration in aging Caenorhabditis elegans. _J. Neurosci._ 34, 3947–3958 (2014). Article CAS PubMed PubMed
Central Google Scholar * Peters, R. Ageing and the brain. _Postgrad. Med J._ 82, 84–88 (2006). Article CAS PubMed PubMed Central Google Scholar * Ye, X., Linton, J. M., Schork, N. J.,
Buck, L. B. & Petrascheck, M. A pharmacological network for lifespan extension in Caenorhabditis elegans. _Aging Cell_ 13, 206–215 (2014). Article CAS PubMed Google Scholar *
Fontana, L., Partridge, L. & Longo, V. D. Extending Healthy Life Span–From Yeast to Humans. _Science_ 328, 321–326 (2010). Article CAS ADS PubMed PubMed Central Google Scholar *
Steinbaugh, M. J., Sun, L. Y., Bartke, A. & Miller, R. A. Activation of genes involved in xenobiotic metabolism is a shared signature of mouse models with extended lifespan. _Am. J.
Physiol. Endocrinol. Metab._ 303, E488–E495 (2012). Article CAS PubMed PubMed Central Google Scholar * Swindell, W. R. Genes and gene expression modules associated with caloric
restriction and aging in the laboratory mouse. _BMC Genomics._ 10, 585 (2009). Article CAS PubMed PubMed Central Google Scholar * Petalcorin, M. I., Joshua, G. W., Agapow, P. M. &
Dolphin, C. T. The fmo genes of Caenorhabditis elegans and C. briggsae: characterisation, gene expression and comparative genomic analysis. _Gene_ 346, 83–96 (2005). Article CAS PubMed
Google Scholar * Beydoun, S. et al. An alternative food source for metabolism and longevity studies in Caenorhabditis elegans. _Commun Biol_. 4, 258
https://doi.org/10.1038/s42003-021-01764-4 (2021). * Bargmann, C. I., Hartwieg, E. & Horvitz, H. R. Odorant-selective genes and neurons mediate olfaction in C. elegans. _Cell_ 74,
515–527 (1993). Article CAS PubMed Google Scholar * Worthy, S. E. et al. Identification of attractive odorants released by preferred bacterial food found in the natural habitats of C.
elegans. _PLoS One_ 13, e0201158 (2018). Article CAS PubMed PubMed Central Google Scholar * Nuttley, W. M., Atkinson-Leadbeater, K. P. & Van Der Kooy, D. Serotonin mediates
food-odor associative learning in the nematode Caenorhabditiselegans. _Proc. Natl Acad. Sci. Usa._ 99, 12449–12454 (2002). Article CAS ADS PubMed PubMed Central Google Scholar *
Flavell, S. W. et al. Serotonin and the neuropeptide PDF initiate and extend opposing behavioral states in C. elegans. _Cell_ 154, 1023–1035 (2013). Article CAS PubMed PubMed Central
Google Scholar * Ro, J. et al. Serotonin signaling mediates protein valuation and aging. E_life_. https://doi.org/10.7554/eLife.16843 (2016) * Linford, N. J., Ro, J., Chung, B. Y. &
Pletcher, S. D. Gustatory and metabolic perception of nutrient stress in Drosophila. _Proc. Natl Acad. Sci. Usa._ 112, 2587–2592 (2015). Article CAS ADS PubMed PubMed Central Google
Scholar * Petrascheck, M., Ye, X. & Buck, L. B. An antidepressant that extends lifespan in adult Caenorhabditis elegans. _Nature_ 450, 553–556 (2007). Article CAS ADS PubMed Google
Scholar * Marshall, R. J. The pharmacology of mianserin-an update. _Br. J. Clin. Pharmacol._ 15, 263S–268S (1983). Article PubMed PubMed Central Google Scholar * Feinberg, S. M.,
Fariba, K. A. & Saadabadi, A. _Thioridazine._ In: StatPearls (StatPearls, 2020). * Zarse, K. & Ristow, M. Antidepressants of the serotonin-antagonist type increase body fat and
decrease lifespan of adult Caenorhabditis elegans. _PLoS One._ 3, e4062 (2008). Article ADS CAS PubMed PubMed Central Google Scholar * Richmond, J. E., Davis, W. S. & Jorgensen, E.
M. UNC-13 is required for synaptic vesicle fusion in C. elegans. _Nat. Neurosci._ 2, 959–964 (1999). Article CAS PubMed PubMed Central Google Scholar * Sieburth, D., Madison, J. M.
& Kaplan, J. M. PKC-1 regulates secretion of neuropeptides. _Nat. Neurosci._ 10, 49–57 (2007). Article CAS PubMed Google Scholar * Bishop, N. A. & Guarente, L. Two neurons
mediate diet-restriction-induced longevity in C. elegans. _Nature_ 447, 545–549 (2007). Article CAS ADS PubMed Google Scholar * Shaw, W. M., Luo, S., Landis, J., Ashraf, J. &
Murphy, C. T. The C. elegans TGF-beta Dauer pathway regulates longevity via insulin signaling. _Curr. Biol._ 17, 1635–1645 (2007). Article CAS PubMed PubMed Central Google Scholar *
Fletcher, M. & Kim, D. H. Age-Dependent Neuroendocrine Signaling from Sensory Neurons Modulates the Effect of Dietary Restriction on Longevity of Caenorhabditis elegans. _PLoS Genet._
13, e1006544 (2017). Article CAS PubMed PubMed Central Google Scholar * Wes, P. D. & Bargmann, C. I. C. elegans odour discrimination requires asymmetric diversity in olfactory
neurons. _Nature_ 410, 698–701 (2001). Article CAS ADS PubMed Google Scholar * Troemel, E. R., Kimmel, B. E. & Bargmann, C. I. Reprogramming chemotaxis responses: sensory neurons
define olfactory preferences in C. elegans. _Cell_ 91, 161–169 (1997). Article CAS PubMed Google Scholar * Murakami, H. & Murakami, S. Serotonin receptors antagonistically modulate
Caenorhabditis elegans longevity. _Aging Cell._ 6, 483–488 (2007). Article CAS PubMed Google Scholar * Sze, J. Y., Victor, M., Loer, C., Shi, Y. & Ruvkun, G. Food and metabolic
signalling defects in a Caenorhabditis elegans serotonin-synthesis mutant. _Nature_ 403, 560–564 (2000). Article CAS ADS PubMed Google Scholar * Trent, C., Tsung, N. & Horvitz, H.
R. Egg-laying defective mutants of the nematode Caeenorhabditis Elegans. _Genetics_ 104, 619–647 (1983). Article CAS PubMed PubMed Central Google Scholar * Rhoades, J. L. et al. ASICs
Mediate Food Responses in an Enteric Serotonergic Neuron that Controls Foraging Behaviors. _Cell_ 176, 85–97.e14 (2019). 01. Article CAS PubMed Google Scholar * Churgin, M. A.,
McCloskey, R. J., Peters, E. & Fang-Yen, C. Antagonistic Serotonergic and Octopaminergic Neural Circuits Mediate Food-Dependent Locomotory Behavior in Caenorhabditis elegans. _J.
Neurosci._ 37, 7811–7823 (2017). Article CAS PubMed PubMed Central Google Scholar * Sulston, J., Dew, M. & Brenner, S. Dopaminergic neurons in the nematode Caenorhabditis elegans.
_J. Comp. Neurol._ 163, 215–226 (1975). Article CAS PubMed Google Scholar * Sawin, E. R., Ranganathan, R. & Horvitz, H. R. C. elegans locomotory rate is modulated by the environment
through a dopaminergic pathway and by experience through a serotonergic pathway. _Neuron_ 26, 619–631 (2000). Article CAS PubMed Google Scholar * Hardaway, J. A. et al. Glial Expression
of the Caenorhabditis elegans Gene swip-10 Supports Glutamate Dependent Control of Extrasynaptic Dopamine Signaling. _J. Neurosci._ 35, 9409–9423 (2015). Article CAS PubMed PubMed Central
Google Scholar * Petrascheck, M., Ye, X. & Buck, L. B. A high-throughput screen for chemicals that increase the lifespan of Caenorhabditis elegans. _Ann. N. Y Acad. Sci._ 1170,
698–701 (2009). Article ADS PubMed Google Scholar * Lorenzo, R., Onizuka, M., Defrance, M. & Laurent, P. Combining single-cell RNA-sequencing with a molecular atlas unveils new
markers for Caenorhabditis elegans neuron classes. _Nucleic Acids Res._ 48, 7119–7134 (2020). CAS PubMed PubMed Central Google Scholar * Svendsen, C. N. et al. Receptor affinity,
neurochemistry and behavioral characteristics of the enantiomers of thioridazine: evidence for different stereoselectivities at D1 and D2 receptors in rat brain. _Neuropharmacology_ 27,
1117–1124 (1988). Article CAS PubMed Google Scholar * Tegowski, M., Fan, C. & Baldwin, A. S. Thioridazine inhibits self-renewal in breast cancer cells via DRD2-dependent STAT3
inhibition, but induces a G. J. _Biol. Chem._ 293, 15977–15990 (2018). Article CAS Google Scholar * Yong, M. et al. DR2 blocker thioridazine: A promising drug for ovarian cancer therapy.
_Oncol. Lett._ 14, 8171–8177 (2017). PubMed PubMed Central Google Scholar * Beaulieu, J. M. & Gainetdinov, R. R. The physiology, signaling, and pharmacology of dopamine receptors.
_Pharm. Rev._ 63, 182–217 (2011). Article CAS PubMed Google Scholar * Missale, C., Nash, S. R., Robinson, S. W., Jaber, M. & Caron, M. G. Dopamine receptors: from structure to
function. _Physiol. Rev._ 78, 189–225 (1998). Article CAS PubMed Google Scholar * Gong, W., Neill, D. B., Lynn, M. & Justice, J. B. Dopamine D1/D2 agonists injected into nucleus
accumbens and ventral pallidum differentially affect locomotor activity depending on site. _Neuroscience_ 93, 1349–1358 (1999). Article CAS PubMed Google Scholar * Wang, D., Yu, Y., Li,
Y. & Wang, Y. Dopamine receptors antagonistically regulate behavioral choice between conflicting alternatives in C. elegans. _PLoS One._ 9, e115985 (2014). Article ADS CAS PubMed
PubMed Central Google Scholar * Suo, S., Culotti, J. G. & Van Tol, H. H. Dopamine counteracts octopamine signalling in a neural circuit mediating food response in C. elegans. _EMBO J._
28, 2437–2448 (2009). Article CAS PubMed PubMed Central Google Scholar * Suo, S. & Ishiura, S. Dopamine modulates acetylcholine release via octopamine and CREB signaling in
Caenorhabditis elegans. _PLoS One._ 8, e72578 (2013). Article CAS ADS PubMed PubMed Central Google Scholar * Noble, T., Stieglitz, J. & Srinivasan, S. An integrated serotonin and
octopamine neuronal circuit directs the release of an endocrine signal to control C. elegans body fat. _Cell Metab._ 18, 672–684 (2013). Article CAS PubMed Google Scholar * Wragg, R. T.
et al. Tyramine and octopamine independently inhibit serotonin-stimulated aversive behaviors in Caenorhabditis elegans through two novel amine receptors. _J. Neurosci._ 27, 13402–13412
(2007). Article CAS PubMed PubMed Central Google Scholar * Ro, J., Harvanek, Z. M. & Pletcher, S. D. FLIC: high-throughput, continuous analysis of feeding behaviors in Drosophila.
_PLoS One._ 9, e101107 (2014). Article ADS CAS PubMed PubMed Central Google Scholar * Cao, J. et al. Comprehensive single-cell transcriptional profiling of a multicellular organism.
_Science_ 357, 661–667 (2017). 08. Article CAS ADS PubMed PubMed Central Google Scholar * Cermak, N. et al. Whole-organism behavioral profiling reveals a role for dopamine in
state-dependent motor program coupling in. E_life_. https://doi.org/10.7554/eLife.57093 (2020) * Hills, T., Brockie, P. J. & Maricq, A. V. Dopamine and glutamate control area-restricted
search behavior in Caenorhabditis elegans. _J. Neurosci._ 24, 1217–1225 (2004). Article CAS PubMed PubMed Central Google Scholar * Lee, K. S. et al. Serotonin-dependent kinetics of
feeding bursts underlie a graded response to food availability in C. elegans. _Nat. Commun._ 8, 14221 (2017). 02. Article CAS ADS PubMed PubMed Central Google Scholar * Li, Z.,
Ichikawa, J., Dai, J. & Meltzer, H. Y. Aripiprazole, a novel antipsychotic drug, preferentially increases dopamine release in the prefrontal cortex and hippocampus in rat brain. _Eur. J.
Pharm._ 493, 75–83 (2004). Article CAS Google Scholar * Bantick, R. A., De Vries, M. H. & Grasby, P. M. The effect of a 5-HT1A receptor agonist on striatal dopamine release.
_Synapse_ 57, 67–75 (2005). Article CAS PubMed Google Scholar * Bang D., et al. Sub-second Dopamine and Serotonin Signaling in Human Striatum during Perceptual Decision-Making. N_euron_.
https://doi.org/10.1016/j.neuron.2020.09.015 (2020) * Benedetto A., Au C., Avila D. S., Milatovic, D. & Aschner M. Extracellular dopamine potentiates mn-induced oxidative stress,
lifespan reduction, and dopaminergic neurodegeneration in a BLI-3-dependent manner in Caenorhabditis elegans. P_LoS Genet_. https://doi.org/10.1371/journal.pgen.1001084 (2010) * Fitch DHA.
_Introduction to nematode evolution and ecology._ WormBook, ed. The C.elegans Research Community: Wormbook; 2005. * Frokjaer-Jensen, C. et al. Single-copy insertion of transgenes in
Caenorhabditis elegans. 10.1038/ng.248. _Nat. Genet_ 40, 1375–1383, https://www.nature.com/ng/journal/v40/n11/suppinfo/ng.248_S1.html (2008). Article CAS PubMed PubMed Central Google
Scholar * Berkowitz L. A., Knight A. L., Caldwell, G. A. & Caldwell K. A. Generation of stable transgenic C. elegans using microinjection. J _Vis Exp_. 2008;https://doi.org/10.3791/833
* Sutphin, G. L. & Kaeberlein M. Measuring Caenorhabditis elegans life span on solid media. J _Vis Exp_. 2009;1152[pii]10.3791/1152 * Anderson, E. N. et al. C. elegans lifespan extension
by osmotic stress requires FUdR, base excision repair, FOXO, and sirtuins. _Mech. Ageing Dev._ 154(Mar), 30–42 (2016). Article CAS PubMed PubMed Central Google Scholar * Maier, W.,
Adilov, B., Regenass, M. & Alcedo, J. A neuromedin U receptor acts with the sensory system to modulate food type-dependent effects on C. elegans lifespan. _PLoS Biol._ 8, e1000376
(2010). Article CAS PubMed PubMed Central Google Scholar * Pang, S. & Curran, S. P. Adaptive capacity to bacterial diet modulates aging in C. elegans. _Cell Metab._ 19, 221–231
(2014). Article CAS PubMed PubMed Central Google Scholar * Greer, E. L. & Brunet, A. Different dietary restriction regimens extend lifespan by both independent and overlapping
genetic pathways in C. elegans. _Aging Cell_ 8, 113–127 (2009). Article CAS PubMed Google Scholar * Linford, N. J., Bilgir, C., Ro, J. & Pletcher, S. D. Measurement of lifespan in
Drosophila melanogaster. J _Vis Exp_. 2013;https://doi.org/10.3791/50068 * Han, S. K. et al. OASIS 2: online application for survival analysis 2 with features for the analysis of maximal
lifespan and healthspan in aging research. _Oncotarget_ 7, 56147–56152 (2016). Article PubMed PubMed Central Google Scholar Download references ACKNOWLEDGEMENTS We thank Dr. Cori
Bargmann for providing the _tph-1_ cell specific knockout strains in ADF or NSM neurons. We thank Dr. Kaveh Ashrafi for providing pKA805[srh-142p::TPH-1] and pKA807[ceh-2p::TPH-1] constructs
for injections. We thank SunyBiotech for generating some strains by injections for this study. We thank all members of the Leiser laboratory for suggestions and discussions. Research
reported in this publication was supported by NIA R01 AG059583 and the Paul F. Glenn Foundation for Aging Research to S.F.L., and NIA F31060663 and the Rackham Predoctoral Fellowship from
the University of Michigan to H.A.M. AUTHOR INFORMATION Author notes * These authors contributed equally: Hillary A. Miller, Shijiao Huang. AUTHORS AND AFFILIATIONS * Cellular and Molecular
Biology Program, University of Michigan, Ann Arbor, MI, 48109, USA Hillary A. Miller, Angela M. Tuckowski & Allyson S. Munneke * Molecular & Integrative Physiology Department,
University of Michigan, Ann Arbor, MI, 48109, USA Shijiao Huang, Elizabeth S. Dean, Megan L. Schaller, Safa Beydoun, Scott D. Pletcher & Scott F. Leiser * Department of Internal
Medicine, University of Michigan, Ann Arbor, MI, 48109, USA Scott F. Leiser Authors * Hillary A. Miller View author publications You can also search for this author inPubMed Google Scholar *
Shijiao Huang View author publications You can also search for this author inPubMed Google Scholar * Elizabeth S. Dean View author publications You can also search for this author inPubMed
Google Scholar * Megan L. Schaller View author publications You can also search for this author inPubMed Google Scholar * Angela M. Tuckowski View author publications You can also search for
this author inPubMed Google Scholar * Allyson S. Munneke View author publications You can also search for this author inPubMed Google Scholar * Safa Beydoun View author publications You can
also search for this author inPubMed Google Scholar * Scott D. Pletcher View author publications You can also search for this author inPubMed Google Scholar * Scott F. Leiser View author
publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS H.A.M., S.H., and S.F.L. developed the conceptual framework and wrote the manuscript. H.A.M., S.H.,
E.S.D., M.L.S., A.M.T., A.S.M., S.B., and S.F.L. contributed to data collection and analysis. S.D.P contributed to the data interpretation and discussion. H.A.M., S.H., and E.S.D. prepared
the figures and tables. All authors reviewed and approved the manuscript. CORRESPONDING AUTHOR Correspondence to Scott F. Leiser. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare
no competing interests. PEER REVIEW PEER REVIEW INFORMATION _Nature Communications_ thanks Arnab Mukhopadhyay and the anonymous reviewer(s) for their contribution to the peer review of this
work. Peer reviewer reports are available. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional
affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION PEER REVIEW FILE DESCRIPTION OF ADDITIONAL SUPPLEMENTARY FILES SUPPLEMENTARY TABLE 1 SUPPLEMENTARY TABLE 2 SUPPLEMENTARY
TABLE 3 SOURCE DATA SOURCE DATA RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing,
adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons
license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a
credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted
use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT
THIS ARTICLE CITE THIS ARTICLE Miller, H.A., Huang, S., Dean, E.S. _et al._ Serotonin and dopamine modulate aging in response to food odor and availability. _Nat Commun_ 13, 3271 (2022).
https://doi.org/10.1038/s41467-022-30869-5 Download citation * Received: 08 March 2021 * Accepted: 12 May 2022 * Published: 07 June 2022 * DOI: https://doi.org/10.1038/s41467-022-30869-5
SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to
clipboard Provided by the Springer Nature SharedIt content-sharing initiative