Rare heterozygous gdf6 variants in patients with renal anomalies
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT Although over 50 genes are known to cause renal malformation if mutated, the underlying genetic basis, most easily identified in syndromic cases, remains unsolved in most patients.
In search of novel causative genes, whole-exome sequencing in a patient with renal, i.e., crossed fused renal ectopia, and extrarenal, i.e., skeletal, eye, and ear, malformations yielded a
rare heterozygous variant in the _GDF6_ gene encoding growth differentiation factor 6, a member of the BMP family of ligands. Previously, _GDF6_ variants were reported to cause pleiotropic
defects including skeletal, e.g., vertebral, carpal, tarsal fusions, and ocular, e.g., microphthalmia and coloboma, phenotypes. To assess the role of _GDF6_ in the pathogenesis of renal
malformation, we performed targeted sequencing in 193 further patients identifying rare _GDF6_ variants in two cases with kidney hypodysplasia and extrarenal manifestations. During
development, _gdf6_ was expressed in the pronephric tubule of _Xenopus laevis_, and _Gdf6_ expression was observed in the ureteric tree of the murine kidney by RNA in situ hybridization.
CRISPR/Cas9-derived knockout of _Gdf6_ attenuated migration of murine IMCD3 cells, an effect rescued by expression of wild-type but not mutant _GDF6_, indicating affected variant function
regarding a fundamental developmental process. Knockdown of _gdf6_ in _Xenopus laevis_ resulted in impaired pronephros development. Altogether, we identified rare heterozygous _GDF6_
variants in 1.6% of all renal anomaly patients and 5.4% of renal anomaly patients additionally manifesting skeletal, ocular, or auricular abnormalities, adding renal hypodysplasia and fusion
to the phenotype spectrum of _GDF6_ variant carriers and suggesting an involvement of GDF6 in nephrogenesis. SIMILAR CONTENT BEING VIEWED BY OTHERS HAPLOINSUFFICIENCY OF _SF3B2_ CAUSES
CRANIOFACIAL MICROSOMIA Article Open access 03 August 2021 HETEROZYGOUS VARIANTS IN THE TEASHIRT ZINC FINGER HOMEOBOX 3 (_TSHZ3_) GENE IN HUMAN CONGENITAL ANOMALIES OF THE KIDNEY AND URINARY
TRACT Article Open access 17 October 2024 EXPANDING THE PHENOTYPIC SPECTRUM AND CLINICAL SEVERITY ASSOCIATED WITH _WLS_ GENE Article 28 April 2023 INTRODUCTION Structural defects of the
kidney range from renal agenesis, hypoplasia, and dysplasia to duplication and fusion phenotypes, such as horseshoe kidneys and crossed fused renal ectopia. The latter is a rare form of
renal anomaly where two fused kidneys come to lie on the same side of the spine, each with their own ureter, one of which crossing the midline to enter the bladder on the contralateral side.
As other renal anomalies are also frequently associated with malformations of the urinary tract, such as ureteropelvic or ureterovesical junction obstruction with hydroureter, or
vesicoureteral reflux (VUR), the term congenital anomalies of the kidney and urinary tract (CAKUT) has been coined to subsume these abnormalities. Taken together, CAKUT phenotypes account
for 15–30% of all prenatally detected congenital malformations [1], and cause around 40% of cases with end-stage kidney disease in children and adolescents [2], thus representing a
significant health burden. In around 85% of patients, CAKUT occur sporadically, while in the remaining 15% of cases familial occurrence is observed. CAKUT may occur in isolation or be part
of a mild or complex syndromal disease. Since over 500 syndromes have been associated with CAKUT [3], it is not surprising that one-third of patients are additionally affected by extrarenal
manifestations [4], and that around 20% of patients may have a genetic disorder that is not detected based on standard clinical evaluation [5]. Although over 50 genes are known to cause
CAKUT in humans if mutated [6, 7], <20% of CAKUT manifestations can be explained by aberrations in these genes [8, 9], indicating a high genetic heterogeneity underlying these defects and
making clear the need to identify new genes associated with renal development and malformation. However, the identification of causative genetic variants in cohorts of sporadic CAKUT
patients and CAKUT families is hampered by variable expressivity, meaning that individuals harboring the same variant may have very different phenotypes within the broad CAKUT spectrum, and
by incomplete penetrance implying that variant carriers may exist that are not affected by a CAKUT phenotype at all. In the last few years, whole-exome sequencing (WES) using next generation
sequencing (NGS) techniques was successfully applied to the study of germline variation underlying human CAKUT [10]. Thereby, novel CAKUT-associated genes were identified e.g., by using a
linkage-based strategy in large CAKUT families [11, 12], a double hit-based strategy in smaller CAKUT families [13], an overlapping strategy in a cohort of patients with similar phenotypes
[14], and a trio-based de novo strategy in patients with sporadic CAKUT [15, 16]. In addition, WES has improved the diagnostic yield of genetic CAKUT causes, particularly in syndromic cases
[9]. Here, in an effort to identify new genes associated with renal malformation in humans, we used a WES approach to determine the genetic variation underlying renal anomalies in an index
patient with syndromic CAKUT. By a targeted mutational screen in a cohort of 193 further patients with renal anomalies, expression analyses, and functional studies in a cellular system
modified by CRISPR/Cas9 genome engineering and an animal model, we suggest that the candidate gene identified in the index patient, _GDF6_, plays a role in kidney development and
malformation. SUBJECTS AND METHODS PATIENTS This study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Boards of Hannover Medical School, Hannover,
Germany, Oslo University Hospital, Oslo, Norway, and University Children’s Hospital, Skopje, North Macedonia. Each family provided informed consent for participation in the study. A total of
194 patients with renal anomalies comprising 122 males and 72 females with a mean age of 10 years (range 1–35 years) were analyzed. Renal phenotypes of all 194 patients are listed in
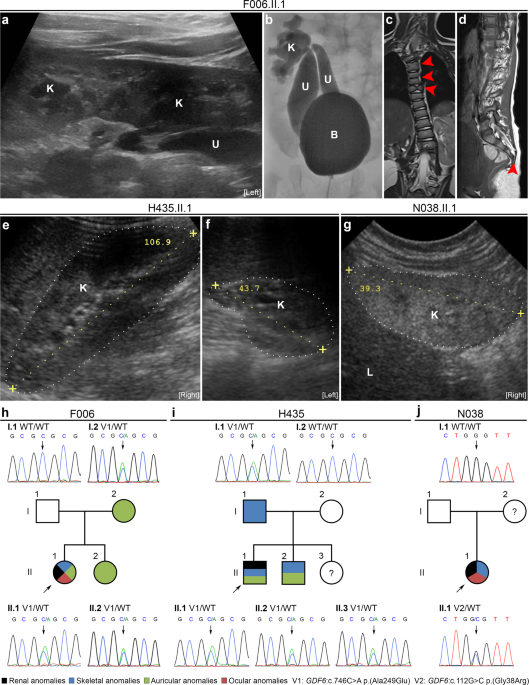
Supplementary Table 1. Rare heterozygous _GDF6_ variants predicted to be disease causing were detected in families F006, H435, and N038. FAMILY F006 The index patient, F006.II.1, born as the
second daughter of non-consanguineous German parents is now 3 years old. After birth, renal ultrasound was notable for left-sided crossed fused renal ectopia (Fig. 1a). Voiding
cystourethrography revealed two megaureters, both with orthotopic ostia in the bladder, one connected to the superior pelvis, and the other to the inferior pelvis of the left-sided fused
kidneys (Fig. 1b), and grade-IV VUR in both ureters. Recurrent urinary tract infections were diagnosed. The patient also showed a left-convex torsion scoliosis, malformations of multiple
vertebral bodies of the cervical and thoracic spine including butterfly and fused vertebrae, and a missing fifth sacral vertebral body and coccyx (Fig. 1c, d). A tethered cord was diagnosed
because of a low standing _conus medullaris_, and detethering surgery was performed. The patient also presented with anal atresia and a rectovestibular fistula, which was surgically
corrected. By echocardiography, two small muscular ventricular septal defects and a patent _foramen ovale_ were diagnosed. Ophthalmologic examination revealed anisometropia with hyperopia,
astigmatism, amblyopia, suspected microphthalmia, corneal opacities, and a best-corrected visual acuity of 0.16 in the left eye and of 1.0 in the right eye. In addition, the patient
presented with a left-sided auricle dysplasia and aplasia of the external auditory canal, while the cochleae and the semicircular canals were unremarkable on both sides according to cranial
MRI. Neurological examination was normal and no developmental or intellectual deficits were observed. Her 6-year-old sister and 36-year-old mother presented with right-sided preauricular
pits, while renal ultrasound examinations revealed no abnormalities. Her father was clinically unremarkable and renal sonography was normal. FAMILY H435 Twelve-year-old patient H435.II.1 was
born as the second son of non-consanguineous Macedonian parents. After birth, the boy presented with a right-sided normal kidney and a left-sided hypodysplastic kidney, a diagnosis
confirmed by ultrasound at age 11 years (Fig. 1e, f). The left kidney could not be visualized by a radionuclide scan using DMSA indicating that it is not functional. In addition, the patient
showed a mild torsion scoliosis. The philtrum appeared shorter than usual, a high-arched palate and malocclusion due to prognathism were noted. His ears were of normal size but had a slight
lop deformity. Except for the presence of a short frenulum, external genitalia were unremarkable. No developmental or intellectual deficits were noted. His 15-year-old brother also had
high-arched palate and lop ears, whereby particularly the right ear was smaller and showed a poorly developed antihelix. A triangular-shaped chin was observed. Neurological examination was
normal and no developmental or intellectual deficits were diagnosed. His 1-year-old sister was clinically unremarkable, but renal ultrasound or other examinations were not performed. His
father presented with short stature. Renal sonography of both parents was normal. FAMILY N038 Fourteen-year-old patient N038.II.1 is the only daughter of an African mother and a Caucasian
father. She was prenatally diagnosed with oligohydramnios and small kidneys. Renal ultrasound at age 6 months showed bilateral renal hypodysplasia as indicated by reduced size,
hyperechogenicity, and reduced corticomedullary differentiation of the kidneys (right kidney shown in Fig. 1g). After birth, she additionally presented with macrocephaly, high-arched palate,
and short narrow palpebral fissures. She required kidney transplantation at the age of 5 years and was re-transplanted at 9 years of age due to chronic rejection and noncompliance with drug
treatment. At 14 years, she has poor kidney function and several comorbidities such as obesity and hypertension. ANIMALS Husbandry and treatment of _Xenopus laevis_ were approved by the
Regierungspräsidium Freiburg, Germany. Mice were kept in accordance with the National Institutes of Health guidelines for the care and use of laboratory animals. All experiments on mice were
approved by the Ethics Board of the Lower Saxony State Office for Consumer Protection and Food Safety. Murine embryos for gene expression analysis were derived from matings of wild-type
mice with NMRI background. For timed pregnancies, vaginal plugs were checked in the morning after mating, and noon was defined as embryonic day (E) 0.5. Embryos were dissected in
phosphate-buffered saline (PBS; Merck, Darmstadt, Germany) and fixed in 4% paraformaldehyde in PBS followed by dehydration using increasing methanol concentrations, i.e., incubation in 25%,
50%, and 75% methanol for 1 h each. Fixed embryos were stored in 100% methanol at −20 °C prior to in situ hybridization analyses. WES AND TARGETED _GDF6_ SEQUENCING WES was performed on
whole-blood DNA of one patient–sibling–parents index family, 30 additional patients with renal malformations, and 74 control individuals using the SureSelectXT Human All Exon V4 target
enrichment kit (Agilent, Santa Clara, CA, USA) on a HiSeq 2000 (Illumina, San Diego, CA, USA) sequencer or the SureSelectXT Human All Exon V5+UTRs target enrichment kit (Agilent) on a HiSeq
2500 (Illumina) sequencer. All samples were sequenced to a mean target coverage of >50×. Sequencing data were aligned to the human reference genome (hg19) using the Biomedical Genomics
Workbench (Qiagen, Hilden, Germany). WES data of the index family were annotated and prioritized using Ingenuity Variant Analysis (Qiagen) and our in-house NGS data analysis workflow as
described in “Results” and summarized in Supplementary Tables 2 and 3. Using conventional chain termination protocols and a 3130XL Genetic Analyzer (Thermo Fisher Scientific, Waltham, MA,
USA), all coding exons and adjacent intronic regions of _GDF6_ were analyzed for sequence variants in 163 further patients with kidney anomalies, selected _GDF6_ variants identified by WES
were verified, and familial segregation analysis was done (oligonucleotide sequences are given in Supplementary Table 4). Nucleotide numbering of the identified variants reflects the
nucleotide position in the coding sequence of human _GDF6_ mRNA (https://www.ncbi.nlm.nih.gov/nuccore/NM_001001557.4) (Supplementary Fig. 1). IMMUNOHISTOCHEMISTRY, RNA IN SITU HYBRIDIZATION,
CRISPR/CAS9 GENOME ENGINEERING AND CELLULAR ASSAYS, KNOCKDOWN AND RESCUE EXPERIMENTS IN _XENOPUS LAEVIS_ Procedures are described in “Supplementary materials” (including Supplementary
Tables 5–8). STATISTICAL ANALYSIS Statistical analysis was done using MATLAB and Statistics Toolbox Release 2018b (The MathWorks, Inc., Natick, MA, USA). Student’s _t_-test or Fisher’s exact
test were used, as applicable, and _p_ values are indicated (*_p_ < 0.05, **_p_ < 0.01, and ***_p_ < 0.001). RESULTS USING WES, A RARE HETEROZYGOUS _GDF6_ MISSENSE VARIANT,
C.746C>A P.(ALA249GLU), WAS DETECTED IN THE INDEX PATIENT Under the assumption that NGS techniques are particularly successful in identifying the genetic cause in patients with syndromic
CAKUT, we applied WES to whole-blood DNA of female patient F006.II.1 with a renal malformation (i.e., crossed fused renal ectopia) as well as skeletal (e.g., scoliosis, fused and butterfly
vertebrae) (Fig. 1a–d), auricular (i.e., auricle dysplasia and aplasia of the external auditory canal), ocular (e.g., anisometropia), and other extrarenal anomalies, and of her mother,
father, and sister who were not affected by renal anomalies. As no rare de novo, homozygous, or compound-heterozygous variants predicted to be disease causing could be detected in the
high-quality exome data of patient F006.II.1, variants were prioritized using the strategy summarized in Supplementary Table 2. Prioritization of high-quality variants of patient F006.II.1
by seriousness, rareness, exclusiveness, and localization in genes (_n_ = 207) reported to be mutated in at least one patient with syndromic CAKUT according to our in-house gene list yielded
five rare (minor allele frequency (MAF) ≤ 1%) non-silent variants not present in controls and predicted to be disease causing by at least one prediction tool (MutationTaster, SIFT, or
PolyPhen-2) (Supplementary Tables 2 and 3). One of these five variants, the _GDF6_ variant c.746C>A p.(Ala249Glu), was presumed causative in patient F006.II.1 because it is reported to be
disease causing by the HGMD Professional (v2018.2; Qiagen) and ClinVar (https://www.ncbi.nlm.nih.gov/clinvar/) databases in patients with skeletal and ocular anomalies [17,18,19,20,21,22]
matching the patient’s extrarenal phenotype (Supplementary Table 9). By direct sequencing, the _GDF6_ c.746C>A variant was confirmed to be heterozygous in the patient as well as in her
sister and mother, both affected by the same mild auricular anomaly (Fig. 1h). The c.746C>A variant is located in the second exon of _GDF6_ and occurs with a MAF of 0.001948 in the global
population cohort and a MAF of 0.003783 in the non-Finnish European cohort of the Genome Aggregation Database (gnomAD v2.1.1) (Table 1). The amino acid alanine at position 249 is
evolutionary conserved and located in the propeptide/prodomain of GDF6 (Supplementary Fig. 1). Based on the ACMG/AMP 2015 guidelines [23], we classified the c.746C>A variant as
“pathogenic” (Table 1). HETEROZYGOUS _GDF6_ MISSENSE VARIANTS WERE DETECTED IN A TOTAL OF THREE OF 194 PATIENTS (1.6%) WITH RENAL MALFORMATIONS AND THREE OF 56 PATIENTS (5.4%) WITH RENAL
PLUS SKELETAL, OCULAR OR AURICULAR ANOMALIES Having identified a _GDF6_ variant previously associated with skeletal and ocular anomalies in a patient presenting with these phenotypes and
additionally with a renal malformation, we explored the frequency of rare _GDF6_ variants in a cohort of patients with renal anomalies. A total of 193 additional cases were either analyzed
by WES (30 patients) or targeted sequencing of the _GDF6_ gene (163 patients). By targeted sequencing, we identified the _GDF6_ variant c.746C>A in an additional patient, H435.II.1 from
North Macedonia, presenting with a hypodysplastic left kidney (Fig. 1f, i) that was nonfunctional on DMSA scan, mild skeletal, and auricular phenotypes, i.e., torsion scoliosis, high-arched
palate, and lop ears (Table 1). The _GDF6_ c.746C>A variant was detected in a heterozygous state in the patient, his father who presented with short stature, his brother who had a
high-arched palate and lop ears, and a 1-year-old sister who was not further examined (Fig. 1i). Another rare heterozygous _GDF6_ missense variant, c.112G>C p.(Gly38Arg), was detected by
targeted sequencing in female patient N038.II.1 presenting with bilateral renal hypodysplasia (Fig. 1g) requiring kidney transplantation at age 5 years in addition to skeletal, i.e.,
macrocephaly and high-arched palate, and ocular, i.e., short narrow palpebral fissures, phenotypes (Fig. 1j and Table 1). As the c.112G>C variant was not inherited from her Caucasian
father, it may be a de novo variant or have been inherited from her African mother (Fig. 1j) from whom no DNA sample was available for clarification. Located in the first exon of _GDF6_
(Supplementary Fig. 1), the c.112G>C variant occurs with a MAF of 0.000656 in the global population cohort, a MAF of 0.007016 in the African population cohort, and is not present in the
non-Finnish European cohort of the gnomAD database v2.1.1 (Table 1). While the c.112G>C variant was of “uncertain significance” based on the ACMG/AMP 2015 guidelines [23], the prediction
tools SIFT and PolyPhen-2 rated it as “damaging” or “possibly damaging” (Table 1). The amino acid glycine at position 38 is evolutionary highly conserved. In total, we detected rare
heterozygous _GDF6_ missense variants predicted to be disease causing in 3 of 194 families with renal anomalies (1.6%). Clinical or radiological reevaluation of the skeleton, eye or ear
revealed skeletal, ocular, or auricular abnormalities in 56 of the 194 patients with renal anomalies analyzed. As the three _GDF6_ variant carriers were among these patients, 3 of 56
patients (5.4%) with renal plus skeletal, ocular, or auricular features carried _GDF6_ variants with a disease-causing prediction, a significant frequency increase compared with that in
renal anomaly patients without abnormalities of the skeleton, eye, or ear (0/138, _p_ = 0.0231, Fisher’s exact test). _GDF6_ IS EXPRESSED IN THE INFANT HUMAN KIDNEY, AND IN THE DEVELOPING
MURINE UROGENITAL SYSTEM AND PRONEPHROS OF _XENOPUS LAEVIS_ Having detected rare _GDF6_ variants in patients with renal anomalies, we went on to explore a possible role for _GDF6_ in kidney
development by determining the expression pattern of _GDF6_ in the infant human kidney as well as in _Mus musculus_ and _Xenopus laevis_ during development. GDF6 protein expression was
detected in the human infant kidney, most prominently in proximal tubules, by immunohistochemistry (Fig. 2a). During lower vertebrate development, expression of _gdf6_ was observed in the
area of the pronephros, the embryonic kidney of _Xenopus laevis_, with particular enhancement in the tissue just adjacent to the pronephric tubule at stage 38 by whole-mount RNA in situ
hybridization. Expression at this stage was also detected around the eye vesicle, branchial arches, notochord, and neural tube of _Xenopus laevis_ embryos (Fig. 2b). During murine urogenital
system development, _Gdf6_ mRNA was present in the ureteric tips at E11.5 shortly after onset of metanephros development. At E13.5, expression of _Gdf6_ in the kidney reached its peak and
was found in all compartments, i.e., the ureter and ureteric tips, of the developing ureteric tree. After onset of collecting duct differentiation at E14.5, _Gdf6_ transcript levels
decreased in the kidney, and renal expression was barely detectable at E18.5. At E11.5, _Gdf6_ was also detectable in the mesothelial lining of the abdominal cavity and weakly in the
epithelium of the urogenital sinus, which develops into the bladder. Expression in the bladder urothelium vanished after E14.5. _Gdf6_ mRNA was also found in the nephric duct or
differentiated _vas deferens_ of male embryos at all analyzed stages (Fig. 2c, data partly not shown). KNOCKOUT OF _GDF6_ IN MURINE INNER MEDULLARY COLLECTING DUCT (MIMCD3) CELLS IMPAIRS
CELL MIGRATION, AN EFFECT RESCUED BY EXPRESSION OF WILD-TYPE NOT MUTANT _GDF6_ IN _GDF6_−/− MIMCD3 CELLS Next, we generated an in vitro test system to determine whether _GDF6_ impacts cell
migration, a central process in development, and to assess whether the identified _GDF6_ variants affect this function. Using CRISPR/Cas9 technology, a single guide RNA targeting the first
coding exon of _Gdf6_ was applied to knockout _Gdf6_ in mIMCD3 cells. Two _Gdf6__−/−_ mIMCD3 cell clones with frameshift variants predicted to result in premature stop codons and
nonfunctional proteins were identified, i.e., clone 32 harboring the homozygous _Gdf6_ variant c.377_378delCA p.(Ser126Cysfs*2), and clone 34 containing the biallelic _Gdf6_ variants
c.373_376delAAGT p.(Lys125Glnfs*9) and c.377_378delCA p.(Ser126Cysfs*2) (Supplementary Fig. 2). A mIMCD3 cell clone with no mutational event in _Gdf6_ (clone 2, _Gdf6__+/+_) was also
identified and used as a control (Supplementary Fig. 2). No differences in cell viability were observed when comparing mIMCD3 cells and _Gdf6__+/+_ mIMCD3 cell clone 2 with the _Gdf6__−/−_
mIMCD3 cell clones 32 or 34 using a cell viability assay (Supplementary Fig. 3). A time series analyzing mIMCD3 cell migration in a wound healing assay resulted in a reduction of the
cell-free area by 50% after 8 h (Supplementary Fig. 4). Therefore, in subsequent analyses the cell-free areas were determined and compared at 0 h and 8 h. While migration of mIMCD3 cells and
_Gdf6__+/+_ mIMCD3 cell clone 2 did not differ significantly, migration of _Gdf6__−/−_ mIMCD3 cell clones 32 and 34 was significantly decreased compared with _Gdf6__+/+_ mIMCD3 cell clone 2
(_p_ = 0.002 and _p_ = 0.005, respectively; Fig. 3a), thereby providing evidence that _Gdf6_ knockout impacts cell migration. Migration was also significantly impaired in _Gdf6__+/−_ mIMCD3
cell clone 30 versus _Gdf6__+/+_ mIMCD3 cell clone 2 (_p_ = 0.041; Supplementary Fig. 5), indicating an effect on migration of heterozygously mutated cells, a cellular model for patients
with heterozygous _GDF6_ variants. A significant increase in migration of _Gdf6__−/−_ mIMCD3 cell clone 32 stably transfected with a wild-type _GDF6_ expression construct was detected
compared with _Gdf6__−/−_ mIMCD3 cell clone 32 transfected with empty vector (_p_ = 0.007; Fig. 3b and Supplementary Fig. 6), showing that re-expression of human wild-type _GDF6_ can rescue
the reduced migration. In contrast, migration of _Gdf6__−/−_ mIMCD3 cell clone 32 transfected with constructs expressing _GDF6_ variants c.112G>C or c.746C>A versus _GDF6_ wild-type
was significantly reduced (_p_ = 0.003 and _p_ = 0.008, respectively; Fig. 3b and Supplementary Fig. 6), demonstrating that the identified _GDF6_ variants are not functional in this assay.
MORPHOLINO (MO) KNOCKDOWN OF _GDF6_ IN _XENOPUS LAEVIS_ IMPAIRS PRONEPHROS DEVELOPMENT To explore a possible role of _gdf6_ in renal development in vivo, we performed _gdf6_ knockdown
unilaterally using a specific _gdf6_ antisense MO in _Xenopus laevis_ tadpoles, and analyzed the developing pronephros, constituting the embryonic kidney in lower vertebrates, at stage 39
(Fig. 4). The pronephric area, calculated as log2 ratio of the injected and uninjected side of the tadpole, was significantly reduced after _gdf6_ MO injection compared with control MO
injection (_p_ = 7.6 × 10−13; Fig. 4). This effect was partially rescued by co-injection of _GDF6_ mRNA with _gdf6_ MO that increased the pronephros area significantly compared with
injecting _gdf6_ MO alone (_p_ = 0.035; Fig. 4). Thereby, we demonstrate a role for _gdf6_ in pronephric tubule development in _Xenopus laevis_. DISCUSSION In the present study
systematically investigating a role of _GDF6_ in renal anomalies, we identified rare heterozygous _GDF6_ variants in 1.6% of patients with kidney malformations. Initially, we found _GDF6_ to
be associated with renal malformation by an unbiased screen for germline variation in a patient with renal as well as skeletal, auricular, ocular, and other anomalies. This patient had been
chosen for WES analysis because NGS technologies have been particularly successful in identifying causative genes in syndromic CAKUT [9]. Subsequently, we detected two further renal anomaly
patients with rare heterozygous _GDF6_ variants among the 193 patients additionally analyzed. In line with the extrarenal manifestations of our three renal anomaly patients carrying _GDF6_
variants, variants in _GDF6_ have previously been reported in patients with skeletal phenotypes, i.e., (i) Klippel–Feil syndrome (KFS) with vertebral segmentation defects frequently
associated with scoliosis, rib abnormalities, and Sprengel’s deformity now known as KFS1 [17, 18], (ii) Chiari malformations [17, 24], and (iii) multiple synostoses syndrome including carpal
and tarsal fusions [25,26,27], as well as ocular phenotypes, i.e., (i) microphthalmia [20,21,22], (ii) coloboma [18, 28], (iii) Leber congenital amaurosis or juvenile retinitis pigmentosa
[19], and (iv) glaucoma [29]. By adding three patients with renal anomalies to the two patients previously reported [17, 18], a total of 5 of 86 (5.8%) individuals carrying rare _GDF6_
variants described here and in the literature (Supplementary Table 9) are known to be affected by congenital kidney malformations. This finding suggests that renal sonography is warranted in
patients carrying rare _GDF6_ variants irrespective of the other abnormalities or disorders they may present with. Vice versa, _GDF6_ mutational analysis may be advisable in patients with
renal anomalies, in particular in those cases additionally affected by the skeletal and ocular phenotypes previously associated with rare _GDF6_ variants and also present in the renal
anomaly patients with _GDF6_ variants of this study. Accordingly, the percentage of rare _GDF6_ variants in patients with renal plus bone, eye, or ear abnormalities was 5.4%, significantly
higher than in renal anomaly patients without these extrarenal manifestations. In this context, it is notable that we detected one _GDF6_ variant, the missense variant c.746C>A,
recurrently in renal anomaly patients, resulting in a significant frequency increase compared with the cohort of the 1000 Genomes Project. Previously, the _GDF6_ c.746C>A variant was
reported in patients with KFS-like skeletal anomalies [17, 18, 20], Chiari malformation [24], micro- or anophthalmia [20,21,22], and Leber congenital amaurosis or juvenile retinitis
pigmentosa [19] (Supplementary Table 9), demonstrating that it can be associated with a spectrum of different phenotypes. In line with these findings, we detected skeletal anomalies in four
of seven _GDF6_ c.746C>A variant carriers from two families, and ocular anomalies in one carrier. Similar to kidney malformations that were present in two of the seven _GDF6_ c.746C>A
variant carriers here, penetrance is reduced for these abnormalities. Reduced penetrance is a known feature associated with renal anomalies. This is exemplified by a family with Stickler
syndrome and a heterozygous nonsense variant in the _BMP4_ gene encoding bone morphogenetic protein 4, a ligand related to GDF6, with renal dysplasia in only one of five family members
carrying the _BMP4_ variant [30]. _GDF6_ encodes growth differentiation factor 6, a member of the BMP family within the transforming growth factor beta (TGF-β) superfamily of ligands that
utilize type I and type II transmembrane serine–threonine kinase receptors [31]. Therefore, in _GDF6_ variant carriers reduced penetrance for renal malformations may be explained by the
complexity of TGF-β signaling comprising numerous ligands (around 30), receptors, and downstream interacting proteins [31], suggesting some redundancy. The GDF6 amino acid sequence consists
of three domains. These are a signal peptide, a propeptide/prodomain, and a mature receptor-binding carboxy-terminal TGF-domain that is highly conserved between species, i.e., it is more
than 90% identical in _Xenopus laevis_ and mouse [32]. Interestingly, 74% of the _GDF6_ variants detected in patients with different phenotypes (Supplementary Table 9), including the
recurrent c.746C>A variant and the c.112G>C variant identified in renal anomaly patients here, affect amino acids located in the prodomain of the GDF6 sequence. Although it is less
conserved, the prodomain of TGF-β superfamily ligands is known to regulate the synthesis, extracellular localization, and activity of these proteins [33]. The prodomain of BMP4, another
member of the BMP family of ligands, for instance, is necessary to generate stable BMP4/7 heterodimers with enhanced bioactivity in vivo [34]. Therefore, it is not unexpected that although
the c.746C>A variant affects an amino acid located in the prodomain of GDF6, the activity of variant GDF6 was significantly decreased in a _SOX9_-reporter luciferase assay [18], and
amounts of variant pre-pro-protein and mature ligand were reduced in the media of transfected cells [19] compared with wild-type. Furthermore, we show here that impaired cell migration of
murine _Gdf6_ knockout cells from the renal inner medullary collecting duct is rescued by expression of wild-type _GDF6_ but not of c.746C>A and c.112G>C variants, indicating that both
are hypomorphic variants with respect to cell movements, a fundamental process in development. BMP signaling is highly implicated in embryogenesis including nephrogenesis and development
[35,36,37,38]. CAKUT phenotypes were observed with high penetrance in mice carrying a heterozygous or homozygous knockout for genes encoding BMP ligands such as BMP4 [39], BMP7 [40], and
GDF11 [41]. Pleiotropic defects have been described as a result of _Gdf6_ knockout, knockdown, or variation in mice, zebrafish, or _Xenopus laevis_ including defects in joint, ligament, and
cartilage formation causing carpal and tarsal fusions and coronal craniosynostosis [42], altered tail tendon fascicles [43], shorter lengths of digits and dermal flat bones in the skull
[44], microphthalmia, anophthalmia, and coloboma [20, 45]. Whether urogenital tract anomalies exist has not previously been examined in these animal models. Based on the results from our
expression and functional studies and the finding that other BMP ligands, such as BMP4, regulate the budding site and elongation of the developing mouse ureter [39], we propose that the rare
_GDF6_ variants detected in five renal anomaly patients here and previously [17, 18] may be causally related to their kidney malformation. This proposal is in line with the spectrum of
renal abnormalities seen in _GDF6_ variant carriers that includes renal agenesis [17] and renal hypodysplasia (two cases here), two anomalies also found in patients carrying _BMP4_ variants
[46]. Kidney agenesis or hypoplasia are potentially caused by defects in ureteric budding and branching of the ureteric tree induced by abberant BMP signaling as detected in _Bmp4_
heterozygous knockout mice [39]. It is quite conceivable that such budding and branching defects giving rise to missing or small dysplastic kidneys also occur in human carriers of rare
_GDF6_ variants because the developing ureteric tree expresses _Gdf6_, and migration of collecting duct cells that originate from the ureteric tree is impaired by _Gdf6_ knockout, as shown
here. Similarly, pronephros size was reduced by _gdf6_ knockdown in _Xenopus laevis_. We and others additionally observed kidney abnormalities involving renal fusions, i.e., a horseshoe
kidney [18] and a crossed fused renal ectopia (one case here), in patients with rare _GDF6_ variants, similar to the skeletal fusions seen (Supplementary Table 9). These data imply that BMP
signaling is also linked to skeletal and renal fusions, similar to observations in a conditional _Bmp4_ knockout mouse in which reduced BMP signaling resulted in hindlimb fusion [47].
According to an established view, renal fusions result as a consequence of abnormal renal ascent during embryogenesis [48], possibly connecting GDF6 to the migration of the kidneys as also
suggested by our cell migration studies. In summary, we identified rare heterozygous _GDF6_ variants in 1.6% of all patients of our renal anomaly cohort, and in 5.4% of those patients
additionally manifesting skeletal, ocular, or auricular abnormalities suggesting that _GDF6_ is associated with human kidney malformations. The phenotype spectrum identified in renal anomaly
patients with _GDF6_ variants ranged from hypodysplasia to fusion. Furthermore, _Gdf6_ expression in the murine developing ureteric tree, diminished migration of murine _Gdf6__−/−_
collecting duct cells and impaired pronephros development after _gdf6_ knockdown in _Xenopus laevis_ may support a role of GDF6 in kidney development. REFERENCES * Queisser-Luft A, Stolz G,
Wiesel A, Schlaefer K, Spranger J. Malformations in newborn: results based on 30,940 infants and fetuses from the Mainz congenital birth defect monitoring system (1990–1998). Arch Gynecol
Obstet. 2002;266:163–7. Article CAS PubMed Google Scholar * Harambat J, van Stralen KJ, Kim JJ, Tizard EJ. Epidemiology of chronic kidney disease in children. Pediatr Nephrol.
2012;27:363–73. Article PubMed Google Scholar * Limwongse C. Syndromes and malformations of the urinary tract. In: Avner E, Harmon W, Niaudet P, Yoshikawa N, editors. Pediatric
nephrology. Berlin, Heidelberg, Germany: Springer; 2009. p. 121–56. Chapter Google Scholar * Stoll C, Dott B, Alembik Y, Roth MP. Associated nonurinary congenital anomalies among infants
with congenital anomalies of kidney and urinary tract (CAKUT). Eur J Med Genet. 2014;57:322–8. Article PubMed Google Scholar * Sanna-Cherchi S, Westland R, Ghiggeri GM, Gharavi AG.
Genetic basis of human congenital anomalies of the kidney and urinary tract. J Clin Invest. 2018;128:4–15. Article PubMed PubMed Central Google Scholar * Kosfeld A, Martens H, Hennies I,
Haffner D, Weber RG. Kongenitale Anomalien der Nieren und ableitenden Harnwege (CAKUT). Med Genet. 2018;30:448–60. CAS Google Scholar * van der Ven AT, Vivante A, Hildebrandt F. Novel
insights into the pathogenesis of monogenic congenital anomalies of the kidney and urinary tract. J Am Soc Nephrol. 2018;29:36–50. Article PubMed Google Scholar * Heidet L, Moriniere V,
Henry C, De Tomasi L, Reilly ML, Humbert C, et al. Targeted exome sequencing identifies PBX1 as involved in monogenic congenital anomalies of the kidney and urinary tract. J Am Soc Nephrol.
2017;28:2901–14. Article CAS PubMed PubMed Central Google Scholar * van der Ven AT, Connaughton DM, Ityel H, Mann N, Nakayama M, Chen J, et al. Whole-exome sequencing identifies
causative mutations in families with congenital anomalies of the kidney and urinary tract. J Am Soc Nephrol. 2018;29:2348–61. Article PubMed PubMed Central Google Scholar * Nigam A,
Knoers N, Renkema KY. Impact of next generation sequencing on our understanding of CAKUT. Semin Cell Dev Biol. 2019;91:104–10. Article CAS PubMed Google Scholar * Vivante A, Kleppa MJ,
Schulz J, Kohl S, Sharma A, Chen J, et al. Mutations in TBX18 cause dominant urinary tract malformations via transcriptional dysregulation of ureter development. Am J Hum Genet.
2015;97:291–301. Article CAS PubMed PubMed Central Google Scholar * Brophy PD, Rasmussen M, Parida M, Bonde G, Darbro BW, Hong X, et al. A gene implicated in activation of retinoic acid
receptor targets is a novel renal agenesis gene in humans. Genetics. 2017;207:215–28. Article CAS PubMed PubMed Central Google Scholar * Humbert C, Silbermann F, Morar B, Parisot M,
Zarhrate M, Masson C, et al. Integrin alpha 8 recessive mutations are responsible for bilateral renal agenesis in humans. Am J Hum Genet. 2014;94:288–94. Article CAS PubMed PubMed Central
Google Scholar * De Tomasi L, David P, Humbert C, Silbermann F, Arrondel C, Tores F, et al. Mutations in GREB1L cause bilateral kidney agenesis in humans and mice. Am J Hum Genet.
2017;101:803–14. Article PubMed PubMed Central CAS Google Scholar * Kosfeld A, Kreuzer M, Daniel C, Brand F, Schafer AK, Chadt A, et al. Whole-exome sequencing identifies mutations of
TBC1D1 encoding a Rab-GTPase-activating protein in patients with congenital anomalies of the kidneys and urinary tract (CAKUT). Hum Genet. 2016;135:69–87. Article CAS PubMed Google
Scholar * Kosfeld A, Brand F, Weiss AC, Kreuzer M, Goerk M, Martens H, et al. Mutations in the leukemia inhibitory factor receptor (LIFR) gene and Lifr deficiency cause urinary tract
malformations. Hum Mol Genet. 2017;26:1716–31. Article CAS PubMed Google Scholar * Tassabehji M, Fang ZM, Hilton EN, McGaughran J, Zhao Z, Bock CEd, et al. Mutations in GDF6 are
associated with vertebral segmentation defects in Klippel–Feil syndrome. Hum Mutat. 2008;29:1017–27. Article CAS PubMed Google Scholar * Asai-Coakwell M, French CR, Ye M, Garcha K, Bigot
K, Perera AG, et al. Incomplete penetrance and phenotypic variability characterize Gdf6-attributable oculo-skeletal phenotypes. Hum Mol Genet. 2009;18:1110–21. Article CAS PubMed Google
Scholar * Asai-Coakwell M, March L, Dai XH, Duval M, Lopez I, French CR, et al. Contribution of growth differentiation factor 6-dependent cell survival to early-onset retinal dystrophies.
Hum Mol Genet. 2013;22:1432–42. Article CAS PubMed Google Scholar * den Hollander AI, Biyanwila J, Kovach P, Bardakjian T, Traboulsi EI, Ragge NK, et al. Genetic defects of GDF6 in the
zebrafish out of sight mutant and in human eye developmental anomalies. BMC Genet. 2010;11:102. Article CAS Google Scholar * Gonzalez-Rodriguez J, Pelcastre EL, Tovilla-Canales JL,
Garcia-Ortiz JE, Amato-Almanza M, Villanueva-Mendoza C, et al. Mutational screening of CHX10, GDF6, OTX2, RAX and SOX2 genes in 50 unrelated microphthalmia-anophthalmia-coloboma (MAC)
spectrum cases. Br J Ophthalmol. 2010;94:1100–4. Article CAS PubMed Google Scholar * Slavotinek AM, Garcia ST, Chandratillake G, Bardakjian T, Ullah E, Wu D, et al. Exome sequencing in
32 patients with anophthalmia/microphthalmia and developmental eye defects. Clin Genet. 2015;88:468–73. Article CAS PubMed PubMed Central Google Scholar * Richards S, Aziz N, Bale S,
Bick D, Das S, Gastier-Foster J, et al. Standards and guidelines for the interpretation of sequence variants: a joint consensus recommendation of the American College of Medical Genetics and
Genomics and the Association for Molecular Pathology. Genet Med. 2015;17:405–24. Article PubMed PubMed Central Google Scholar * Markunas CA, Soldano K, Dunlap K, Cope H, Asiimwe E,
Stajich J, et al. Stratified whole genome linkage analysis of Chiari type I malformation implicates known Klippel-Feil syndrome genes as putative disease candidates. PLoS ONE. 2013;8:e61521.
Article CAS PubMed PubMed Central Google Scholar * Drage Berentsen R, Haukanes BI, Juliusson PB, Rosendahl K, Houge G. A novel GDF6 mutation in a family with multiple synostoses
syndrome without hearing loss. Mol Syndromol. 2019;9:228–34. Article PubMed CAS Google Scholar * Terhal PA, Verbeek NE, Knoers N, Nievelstein R, van den Ouweland A, Sakkers RJ, et al.
Further delineation of the GDF6 related multiple synostoses syndrome. Am J Med Genet A. 2018;176:225–9. Article CAS PubMed Google Scholar * Wang J, Yu T, Wang Z, Ohte S, Yao R-E, Zheng
Z, et al. A new subtype of multiple synostoses syndrome is caused by a mutation in GDF6 that decreases its sensitivity to noggin and enhances its potency as a BMP signal. J Bone Min Res.
2016;31:882–9. Article CAS Google Scholar * Chassaing N, Causse A, Vigouroux A, Delahaye A, Alessandri J-L, Boespflug-Tanguy O, et al. Molecular findings and clinical data in a cohort of
150 patients with anophthalmia/microphthalmia. Clin Genet. 2014;86:326–34. Article CAS PubMed Google Scholar * Huang X, Xiao X, Jia X, Li S, Li M, Guo X, et al. Mutation analysis of the
genes associated with anterior segment dysgenesis, microcornea and microphthalmia in 257 patients with glaucoma. Int J Mol Med. 2015;36:1111–7. Article CAS PubMed Google Scholar * Nixon
TRW, Richards A, Towns LK, Fuller G, Abbs S, Alexander P, et al. Bone morphogenetic protein 4 (BMP4) loss-of-function variant associated with autosomal dominant Stickler syndrome and renal
dysplasia. Eur J Hum Genet. 2019;27:369–77. Article CAS PubMed Google Scholar * De Caestecker M. The transforming growth factor-b: superfamily of receptors. Cytokine Growth Factor Rev.
2004;15:1–11. Article CAS PubMed Google Scholar * Chang C, Hemmati-Brivanlou A. Xenopus GDF6, a new antagonist of noggin and a partner of BMPs. Development. 1999;126:3347–57. Article
CAS PubMed Google Scholar * Harrison CA, Al-Musawi SL, Walton KL. Prodomains regulate the synthesis, extracellular localisation and activity of TGF-β superfamily ligands. Growth Factors.
2011;29:174–86. Article CAS PubMed Google Scholar * Neugebauer JM, Kwon S, Kim HS, Donley N, Tilak A, Sopory S, et al. The prodomain of BMP4 is necessary and sufficient to generate
stable BMP4/7 heterodimers with enhanced bioactivity in vivo. Proc Natl Acad Sci USA. 2015;112:E2307–16. Article CAS PubMed PubMed Central Google Scholar * Cain JE, Hartwig S, Bertram
JF, Rosenblum ND. Bone morphogenetic protein signaling in the developing kidney: present and future. Differentiation. 2008;76:831–42. Article CAS PubMed Google Scholar * Nishinakamura R,
Sakaguchi M. BMP signaling and its modifiers in kidney development. Pediatr Nephrol. 2014;29:681–6. Article PubMed Google Scholar * Wang RN, Green J, Wang Z, Deng Y, Qiao M, Peabody M,
et al. Bone morphogenetic protein (BMP) signaling in development and human diseases. Genes Dis. 2014;1:87–105. Article CAS PubMed PubMed Central Google Scholar * Mamo TM, Wittern AB,
Kleppa M-J, Bohnenpoll T, Weiss A-C, Kispert A. BMP4 uses several different effector pathways to regulate proliferation and differentiation in the epithelial and mesenchymal tissue
compartments of the developing mouse ureter. Hum Mol Genet. 2017;26:3553–63. Article CAS PubMed Google Scholar * Miyazaki Y, Oshima K, Fogo A, Hogan BL, Ichikawa I. Bone morphogenetic
protein 4 regulates the budding site and elongation of the mouse ureter. J Clin Invest. 2000;105:863–73. Article CAS PubMed PubMed Central Google Scholar * Dudley AT, Lyons KM,
Robertson EJ. A requirement for bone morphogenetic protein-7 during development of the mammalian kidney and eye. Genes Dev. 1995;9:2795–807. Article CAS PubMed Google Scholar * Esquela
AF, Lee SJ. Regulation of metanephric kidney development by growth/differentiation factor 11. Dev Biol. 2003;257:356–70. Article CAS PubMed Google Scholar * Settle SH, Rountree RB, Sinha
A, Thacker A, Higgins K, Kingsley DM. Multiple joint and skeletal patterning defects caused by single and double mutations in the mouse Gdf6 and Gdf5 genes. Dev Biol. 2003;254:116–30.
Article CAS PubMed Google Scholar * Mikic B, Rossmeier K, Bierwert L. Identification of a tendon phenotype in GDF6 deficient mice. Anat Rec. 2009;292:396–400. Article Google Scholar *
Indjeian VB, Kingman GA, Jones FC, Guenther CA, Grimwood J, Schmutz J, et al. Evolving new skeletal traits by cis-regulatory changes in bone morphogenetic proteins. Cell. 2016;164:45–56.
Article CAS PubMed PubMed Central Google Scholar * Asai-Coakwell M, French CR, Berry KM, Ye M, Koss R, Somerville M, et al. GDF6, a novel locus for a spectrum of ocular developmental
anomalies. Am J Hum Genet. 2007;80:306–15. Article CAS PubMed Google Scholar * Weber S, Taylor JC, Winyard P, Baker KF, Sullivan-Brown J, Schild R, et al. SIX2 and BMP4 mutations
associate with anomalous kidney development. J Am Soc Nephrol. 2008;19:891–903. Article CAS PubMed PubMed Central Google Scholar * Suzuki K, Adachi Y, Numata T, Nakada S, Yanagita M,
Nakagata N, et al. Reduced BMP signaling results in hindlimb fusion with lethal pelvic/urogenital organ aplasia: a new mouse model of sirenomelia. PLoS ONE. 2012;7:e43453. Article CAS
PubMed PubMed Central Google Scholar * Taghavi K, Kirkpatrick J, Mirjalili SA. The horseshoe kidney: Surgical anatomy and embryology. J Pediatr Urol. 2016;12:275–80. Article CAS PubMed
Google Scholar Download references ACKNOWLEDGEMENTS We deeply appreciate the participation of patients and families in this study. We wish to thank Achim Gossler and Michael Klintschar,
Hannover Medical School, Hannover, Germany for providing mIMCD3 cells and human infant kidney sections, respectively. The _Xenopus laevis gdf6_ construct for whole-mount RNA in situ
hybridization was kindly provided by Alexandra Schambony, Friedrich-Alexander University Erlangen-Nuremberg, Erlangen, Germany. We wish to acknowledge the assistance of the Cell Sorting Core
Facility of Hannover Medical School. Open Access Funding provided by Projekt DEAL. FUNDING This research was supported by grants from the Else Kröner-Fresenius-Stiftung to RGW (2014_A234),
the Deutsche Forschungsgemeinschaft to AnC (KO5614/2-1) and SSL (LI1817/2-1), the Swiss National Science Foundation (SNF) to SSL (NCCR Kidney.CH), and the Hochschulinterne Leistungsförderung
(HiLF) program of Hannover Medical School to FB. The Cell Sorting Core Facility of Hannover Medical School is supported by the Braukmann-Wittenberg-Herz-Stiftung and the Deutsche
Forschungsgemeinschaft. AUTHOR INFORMATION Author notes * These authors contributed equally: Dieter Haffner, Ruthild G. Weber AUTHORS AND AFFILIATIONS * Department of Human Genetics,
Hannover Medical School, 30625, Hannover, Germany Helge Martens, Anne Christians, Frank Brand & Ruthild G. Weber * Department of Pediatric Kidney, Liver and Metabolic Diseases, Hannover
Medical School, 30625, Hannover, Germany Imke Hennies & Dieter Haffner * Department of Medicine, Renal Division, University Medical Center Freiburg, Faculty of Medicine, University of
Freiburg, 79110, Freiburg, Germany Maike Getwan & Soeren S. Lienkamp * Institute of Anatomy and Zurich Center for Integrative Human Physiology (ZIHP), University of Zurich, 8057, Zurich,
Switzerland Maike Getwan & Soeren S. Lienkamp * Institute of Molecular Biology, Hannover Medical School, 30625, Hannover, Germany Anna-Carina Weiss & Andreas Kispert * Division of
Paediatric and Adolescent Medicine, Oslo University Hospital, 0424, Oslo, Norway Ann Christin Gjerstad & Anna Bjerre * Department of Neuropathology, Institute of Pathology, Hannover
Medical School, 30625, Hannover, Germany Arne Christians * Medical Faculty Skopje, University Children’s Hospital, 1000, Skopje, North Macedonia Zoran Gucev & Velibor Tasic * Genome
Analytics Research Group, Helmholtz Centre for Infection Research, 38124, Braunschweig, Germany Robert Geffers * Department of Paediatrics and Transplantation Center, University Hospital
Motol, Second Faculty of Medicine, Charles University, 150 06, Prague, Czech Republic Tomáš Seeman Authors * Helge Martens View author publications You can also search for this author
inPubMed Google Scholar * Imke Hennies View author publications You can also search for this author inPubMed Google Scholar * Maike Getwan View author publications You can also search for
this author inPubMed Google Scholar * Anne Christians View author publications You can also search for this author inPubMed Google Scholar * Anna-Carina Weiss View author publications You
can also search for this author inPubMed Google Scholar * Frank Brand View author publications You can also search for this author inPubMed Google Scholar * Ann Christin Gjerstad View author
publications You can also search for this author inPubMed Google Scholar * Arne Christians View author publications You can also search for this author inPubMed Google Scholar * Zoran Gucev
View author publications You can also search for this author inPubMed Google Scholar * Robert Geffers View author publications You can also search for this author inPubMed Google Scholar *
Tomáš Seeman View author publications You can also search for this author inPubMed Google Scholar * Andreas Kispert View author publications You can also search for this author inPubMed
Google Scholar * Velibor Tasic View author publications You can also search for this author inPubMed Google Scholar * Anna Bjerre View author publications You can also search for this author
inPubMed Google Scholar * Soeren S. Lienkamp View author publications You can also search for this author inPubMed Google Scholar * Dieter Haffner View author publications You can also
search for this author inPubMed Google Scholar * Ruthild G. Weber View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS HM, AnC, FB, DH, and RGW
designed research; HM analyzed WES data, performed and provided figures of all experiments except RNA in situ hybridization and _Xenopus_ studies, compiled literature data; HM, AnC, FB, and
RGW analyzed these data; MG and SSL performed, analyzed, and provided figures of _Xenopus_ studies; ACW and AK performed, analyzed, and provided figures of RNA in situ hybridization on
murine tissue; IH, ACG, ZG, TS, VT, AB, and DH performed clinical examinations, contributed patient materials, data, and clinical expertise; ArC provided technical expertise; RG generated
WES data; HM, AnC, FB, DH, and RGW wrote the manuscript; all authors reviewed and revised the manuscript. CORRESPONDING AUTHOR Correspondence to Ruthild G. Weber. ETHICS DECLARATIONS
CONFLICT OF INTEREST The authors declare that they have no conflict of interest. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with regard to jurisdictional claims
in published maps and institutional affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY MATERIAL SUPPLEMENTARY TABLE 3 RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a
Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit
to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are
included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and
your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this
license, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Martens, H., Hennies, I., Getwan, M. _et al._ Rare heterozygous
_GDF6_ variants in patients with renal anomalies. _Eur J Hum Genet_ 28, 1681–1693 (2020). https://doi.org/10.1038/s41431-020-0678-9 Download citation * Received: 20 March 2020 * Revised: 05
June 2020 * Accepted: 15 June 2020 * Published: 31 July 2020 * Issue Date: December 2020 * DOI: https://doi.org/10.1038/s41431-020-0678-9 SHARE THIS ARTICLE Anyone you share the following
link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature
SharedIt content-sharing initiative