Boceprevir, gc-376, and calpain inhibitors ii, xii inhibit sars-cov-2 viral replication by targeting the viral main protease
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT A new coronavirus SARS-CoV-2, also called novel coronavirus 2019 (2019-nCoV), started to circulate among humans around December 2019, and it is now widespread as a global pandemic.
The disease caused by SARS-CoV-2 virus is called COVID-19, which is highly contagious and has an overall mortality rate of 6.35% as of May 26, 2020. There is no vaccine or antiviral
available for SARS-CoV-2. In this study, we report our discovery of inhibitors targeting the SARS-CoV-2 main protease (Mpro). Using the FRET-based enzymatic assay, several inhibitors
including boceprevir, GC-376, and calpain inhibitors II, and XII were identified to have potent activity with single-digit to submicromolar IC50 values in the enzymatic assay. The mechanism
of action of the hits was further characterized using enzyme kinetic studies, thermal shift binding assays, and native mass spectrometry. Significantly, four compounds (boceprevir, GC-376,
calpain inhibitors II and XII) inhibit SARS-CoV-2 viral replication in cell culture with EC50 values ranging from 0.49 to 3.37 µM. Notably, boceprevir, calpain inhibitors II and XII
represent novel chemotypes that are distinct from known substrate-based peptidomimetic Mpro inhibitors. A complex crystal structure of SARS-CoV-2 Mpro with GC-376, determined at 2.15 Å
resolution with three protomers per asymmetric unit, revealed two unique binding configurations, shedding light on the molecular interactions and protein conformational flexibility
underlying substrate and inhibitor binding by Mpro. Overall, the compounds identified herein provide promising starting points for the further development of SARS-CoV-2 therapeutics. SIMILAR
CONTENT BEING VIEWED BY OTHERS BOTH BOCEPREVIR AND GC376 EFFICACIOUSLY INHIBIT SARS-COV-2 BY TARGETING ITS MAIN PROTEASE Article Open access 04 September 2020 IDENTIFICATION OF SARS-COV-2
MPRO INHIBITORS CONTAINING P1’ 4-FLUOROBENZOTHIAZOLE MOIETY HIGHLY ACTIVE AGAINST SARS-COV-2 Article Open access 25 February 2023 DEVELOPMENT OF OPTIMIZED DRUG-LIKE SMALL MOLECULE INHIBITORS
OF THE SARS-COV-2 3CL PROTEASE FOR TREATMENT OF COVID-19 Article Open access 07 April 2022 INTRODUCTION An emerging respiratory disease COVID-19 started to circulate among human in December
2019. Since its first outbreak in China from an unknown origin, it quickly became a global pandemic. As of May 26, 2020, there are 343,562 deaths among 5,406,282 confirmed cases in 215
countries (https://www.who.int/emergencies/diseases/novel-coronavirus-2019). The etiological pathogen of COVID-19 is a new coronavirus, the severe acute respiratory syndrome coronavirus 2
(SARS-CoV-2), also called novel coronavirus (nCoV-2019). As the name indicates, SARS-CoV-2 is similar to SARS-CoV, the virus that causes severe acute respiratory symptoms in human and killed
774 people among 8098 infected worldwide in 2003.1 SARS-CoV-2 shares ~82% of sequence identity with SARS-CoV and to a less extent with Middle East respiratory syndrome coronavirus
(MERS-CoV) (~50%).2,3 SARS-CoV-2 is an enveloped, positive-sense, single-stranded RNA virus that belongs to the β-lineage of the coronavirus.4 The β-lineage also contains two other important
human pathogens, the SARS-CoV and MERS-CoV. The mortality rate of COVID-19 is around 6.35% as of May 26, 2020, which is lower than that of SARS (~10%) and MERS (~34%).1 However, current
data indicate that COVID-19 is more contagious and has a larger R0 value than SARS and MERS5, resulting in higher overall death tolls than SARS and MERS. The SARS-CoV-2 virus is currently
spreading at an alarming speed in Europe and the United States. There is currently no antiviral or vaccine for SARS-CoV-2. The SARS-CoV-2 viral genome encodes a number of structural proteins
(e.g., capsid spike glycoprotein), non-structural proteins (e.g., 3-chymotrypsin-like protease (3CL or main protease), papain-like protease, helicase, and RNA-dependent RNA polymerase), and
accessary proteins. Compounds that target any of these viral proteins might be potential antiviral drug candidates.6,7 In this study, we focus on the viral 3CL protease, also called the
main protease (Mpro), and aim to develop potent Mpro inhibitors as SAR-CoV-2 antivirals. The Mpro plays an essential role in coronavirus replication by digesting the viral polyproteins at
more than 11 sites, and it appears like a high-profile target for antiviral drug discovery.8,9,10,11 The Mpro has a unique substrate preference for glutamine at the P1 site (Leu-Gln↓(Ser,
Ala, Gly)), a feature that is absent in closely related host proteases, suggesting that it is feasible to achieve high selectivity by targeting viral Mpro. As such, we developed the
fluorescence resonance energy transfer (FRET)-based enzymatic assay for the SARS-CoV-2 Mpro and applied it to screen a focused library of protease inhibitors. Here we report our findings of
several novel hits targeting SARS-CoV-2 Mpro and their mechanism of action. The in vitro antiviral activity of the hits was also evaluated in cell culture using infectious SARS-CoV-2 virus.
Overall, our study provides a list of drug candidates for SARS-CoV-2 with a confirmed mechanism of action, and the results might help speed up the drug discovery efforts in combating
COVID-19. The compounds identified herein represent some of the most potent and selective SARS-CoV-2 Mpro inhibitors so far with both enzymatic inhibition and cellular antiviral
activity.8,10,11 The X-ray crystal structure of SARS-CoV-2 Mpro with GC-376 showed that the compound can adopt two configurations R and S, offering a molecular explanation of the high
binding affinity of the aldehyde-containing inhibitors. Significantly, the discovery of calpain II and XII inhibitors as potent SARS-CoV-2 antivirals suggests that it might be feasible to
design dual inhibitors against the viral Mpro and the host calpains/cathepsins, both of which are important for viral replication. RESULTS ESTABLISHING THE FRET-BASED ASSAY FOR THE
SARS-COV-2 MPRO The Mpro gene from SARS-CoV-2 strain BetaCoV/Wuhan/WIV04/2019 was inserted into pET-29a(+) vector and expressed in BL21(DE3) _Escherichia Coli_. with a His-tag in its
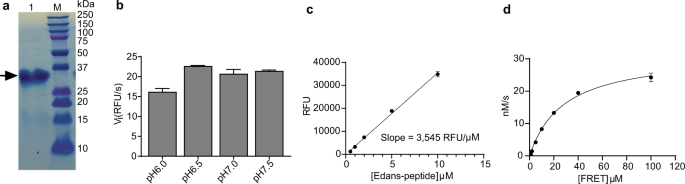
C-terminus. The Mpro protein was purified with Ni-NTA column to high purity (Fig. 1a). The determined molecular weight of Mpro is 34,861 Da from native mass spectrometry, which matches the
calculated molecular weight of Mpro without the N-terminal methionine (Supplementary information, Fig. S1). The first methionine might be cleaved by the methionine aminopeptidase as reported
before.12 To establish the FRET assay condition, we designed a FRET based substrate using the sequence between viral polypeptide NSP4-NSP5 junction from SARS-CoV-2:
Dabcyl-KTSAVLQ/SGFRKME(Edans).8 We then tested the Mpro proteolytic activity in buffers with different pH. We found that Mpro displays the highest activity in pH 6.5 buffer (Fig. 1b), which
contains 20 mM HEPES, 120 mM NaCl, 0.4 mM EDTA, 4 mM 1,4-dithiothreitol (DTT) and 20% glycerol. As such, all the following proteolytic assay was conducted using this pH 6.5 buffer. A
standard curve was generated to convert the relative fluorescence unit (RFU) to the amount of the cleaved substrate (nM) such that kcat can be calculated (Fig. 1c). Next, we characterized
the enzymatic activity of this SARS-CoV-2 Mpro by measuring the _K_m and _V_max values. When 200 nM Mpro was mixed with various concentration of FRET substrate (0–100 µM), the initial
velocity was measured and plotted against substrate concentration. Curve fitting with Michaelis–Menten equation gave the best-fit values of _K_m and _V_max as 28.2 ± 3.4 µM and 31.7 ± 1.6
RFU/s, respectively (Fig. 1d). The calculated _k_cat/_K_m was 5,624 s−1 M−1, which is similar to the previously reported value of 3,426.1 s−1 M−1 by Hilgenfeld et al.8, suggesting that the
C-terminal His-tag Mpro is as active as the native Mpro. PRIMARY SCREENING OF A FOCUSED PROTEASE INHIBITOR LIBRARY AGAINST THE SARS-COV-2 MPRO With the established FRET assay condition, we
screened a collection of protease inhibitors from the Selleckchem bioactive compound library to identify potential SARS-CoV-2 Mpro inhibitors (Table 1). The protease inhibitors are grouped
based on their targets and mechanism of action, including proteasome inhibitors (1–8), HIV protease inhibitors (9–14), γ-secretase inhibitors (15–22), HCV NS3–4A protease inhibitors (23–29),
DPP-4 inhibitors (30–35), miscellaneous serine protease inhibitors (36–39), cathepsin and calpain protease inhibitors (40–43), miscellaneous cysteine protease inhibitors (44–48), matrix
metalloprotease inhibitors (49–51), and miscellaneous protease inhibitors (52–55). The inhibitors were pre-incubated with 100 nM of Mpro at 30 °C for 30 min in the presence of 4 mM DTT
before the addition of 10 µM FRET substrate. The addition of DTT was to quench non-specific thiol reactive compounds and to ensure that Mpro is in the reducing condition. All compounds were
tested at 20 µM, except compound 26, which was tested at 2 µM due to its fluorescent background. Encouragingly, four inhibitors (24, 28, 29 and 43) showed more than 60% inhibition against
Mpro at 20 µM (Fig. 2). Among the hits, simeprevir (24), boceprevir (28), and narlaprevir (29) are HCV NS3–4A serine protease inhibitors, and compound MG-132 (43) inhibits both proteasome
and calpain. SECONDARY SCREENING OF A FOCUSED LIBRARY OF CALPAIN/CATHEPSIN INHIBITORS AND KNOWN VIRAL 3CLPRO INHIBITORS Given the encouraging results from the primary screening, we then
further characterized the four hits (24, 28, 29, and 43) in a combination of assays including dose-response titration, thermal shift binding assay (TSA), and counter screening assays with
two other viral cysteine proteases, the enterovirus A71 (EV-A71) 2A and 3C proteases, both of which are cysteine proteases (Table 2). The HCV NS3–4A protease inhibitors boceprevir (28) and
narlaprevir (29) inhibited Mpro with 50% inhibitory concentration (IC50) values of 4.13 and 4.73 µM, respectively (Table 2), more potent than simeprevir (24) (IC50 = 13.74 µM). Both
compounds (28 and 29) also showed strong binding towards Mpro and shifted the melting temperature of the protein (ΔTm) by 6.67 °C and 5.18 °C, respectively, at 40 µM. Despite their potent
inhibition against the HCV NS3–4A serine protease and the SARS-CoV-2 cysteine Mpro, boceprevir (28) and narlaprevir (29) did not inhibit the EV-A71 2A and 3C proteases (IC50 > 20 µM),
suggesting that they are not non-specific cysteine protease inhibitors. The calpain inhibitor MG-132 (43) had an IC50 value of 3.90 µM against Mpro, and was not active against the EV-A71 2A
and 3C proteases (IC50 > 20 µM). The binding of MG-132 (43) to Mpro was also confirmed in the TSA assay with a ΔTm of 4.02 °C. In light of the promising results of the calpain inhibitor
MG-132 (43), we then pursued testing for other calpain and cathepsin inhibitors that are commercially available (56–63) (Table 2). These compounds were not included in the initial library
because they have not been advanced to clinical studies. Among this series of analogs, calpain inhibitor II (61) and XII (62) are the most potent Mpro inhibitors with IC50 values of 0.97 and
0.45 µM, respectively. Binding of compounds (61 and 62) to Mpro shifted the melting curve of the protein by 6.65 and 7.86 °C, respectively. Encouragingly, both compounds (61 and 62) did not
inhibit the EV-A71 2A and 3C proteases (IC50 > 20 µM). Calpain inhibitor I (59) and MG-115 (60) also showed potent inhibition against Mpro with IC50 values of 8.60 and 3.14 µM,
respectively. Calpeptin (56) and PSI (63) had moderate activity against Mpro with IC50 values of 10.69 and 10.38 µM, respectively. In contrast, calpain inhibitors III (57) and VI (58) were
not active (IC50 > 20 µM). We also included two well-known viral 3CL protease inhibitors GC-376 (64) and rupintrivir (65) in the secondary screening. GC-376 (64) is an investigational
veterinary drug that is being developed for feline infectious peritonitis (FIP).13,14 GC-376 (64) was designed to target the viral 3CL protease and had potent antiviral activity against
multiple viruses including MERS-CoV, FIPV, and norovirus.13,15 Rupintrivir (65) was developed as a rhinovirus antiviral by targeting the viral 3CL protease, but it was discontinued in
clinical trials due to side effects.16 In our study, we found that GC-376 (64) was the most potent Mpro inhibitor with an IC50 value of 0.03 µM. It shifted the melting curve of Mpro by 18.30
°C upon binding. In contrast, rupintrivir (65) was not active against Mpro (IC50 > 20 µM). Previous report also showed that rupintrivir was not active against the SARS-CoV 3CLpro (Mpro)
(IC50 > 100 µM).17 Both compounds (64 and 65) were inactive against the EV-A71 2A protease, but showed potent inhibition against the EV-A71 3C protease, which is consistent with
previously reported results.15,18,19 When plotting the IC50 values (log scale) of the inhibitors against Mpro from the FRET enzymatic assay with the melting temperature shifts (ΔTm) from TSA
(Fig. 3a), a linear correlation was observed, and the r2 of the linear regression fitting is 0.94. This suggests that there is a direct correlation between the enzymatic inhibition and
protein binding: a more potent enzyme inhibitor also binds to the protein with higher affinity. The stabilization of the Mpro against thermal denaturation was also compound concentration
dependent (Fig. 3b). The binding of the four most potent inhibitors boceprevir (28), calpain inhibitors II (61), XII (62), and GC-376 (64) to SARS-CoV-2 Mpro was further characterized by
native mass spectrometry (MS) (Fig. 3c–f). Native MS analysis showed that Mpro formed a dimer complex with a mass of 69,722 Da, indicating that the extra N-terminal methionine in our Mpro
construct was cleaved, yielding the native Mpro N-terminus with a serine residue (Supplementary information, Fig. S1). A small amount of protomer and dimer with a single C-terminal
truncation of the His-tag was also observed, but the intact dimer was the predominant signal (Supplementary information, Fig. S1). Addition of all four ligands tested, boceprevir (28),
calpain inhibitors II (61), XII (62), and GC-376 (64), showed binding of up to two ligands per dimer (Fig. 3c–f), suggesting a binding stoichiometry of one drug per protomer. MECHANISM OF
ACTION OF HITS To elucidate the mechanism of action of hits against SARS-CoV-2 Mpro, we focus on five most potent compounds prioritized from the primary and secondary screenings including
boceprevir (28), MG-132 (43), calpain inhibitor II (61), calpain inhibitor XII (62), and GC-376 (64). For this, we performed enzyme kinetic studies with different concentrations of
inhibitors (Fig. 4). A biphasic enzymatic progression curve in the presence but not absence of inhibitor is typically a hallmark for a slow covalent binding inhibitor. In Fig. 4, the left
column shows the progression curves up to 4 h. Biphasic progression curves were observed for all 5 inhibitors at high drug concentrations. Significant substrate depletion was observed when
the proteolytic reaction proceeded beyond 90 min. We therefore chose the first 90 min of the progression curves for curve fitting (Fig. 4, middle column). We fit the progression curves in
the presence of different concentrations of GC-376 (64) with the two-step Morrison equation (Eq. (3) in “Materials and methods” section). GC-376 (64) binds to SARS-CoV-2 Mpro with an
equilibrium dissociation constant for the inhibitor (KI) of 59.9 ± 21.7 nM in the first step. After initial binding, a covalent bond is formed at a slower velocity between GC-376 (64) and
Mpro with the second reaction rate constant (k2) being 0.00245 ± 0.00047 s−1, resulting in an overall k2/KI value of 4.08 × 104 M−1 s−1 (Fig. 4a). However, when we tried to fit the
proteolytic progression curves for boceprevir (28), MG-132 (43), calpain inhibitors II (61) and XII (62) using the same two-step reaction mechanism, we could not obtain accurate values for
the second rate constant k2. This is presumably due to significant substrate depletion before the equilibrium between EI and EI*, leading to very small values of k2. Accordingly, for these
four inhibitors (28, 43, 61, and 62), only the dissociation constant KI values from the first step were determined (Fig. 4b–e). The inhibition constants (KI) for boceprevir (28), MG-132
(43), calpain inhibitors II (61) and XII (62) are 1.18 ± 0.10 µM, 1.57 ± 0.13 µM, 0.40 ± 0.02 µM, and 0.13 ± 0.02 µM, respectively. CELLULAR ANTIVIRAL ACTIVITY AND CYTOTOXICITY OF HITS To
test the hypothesis that inhibiting the enzymatic activity of Mpro will lead to the inhibition of SARS-CoV-2 viral replication, we performed cellular antiviral assays for the five promising
hits (64, 28, 43, 61, and 62) against SARS-CoV-2. For this, we first tested the cellular cytotoxicity of these compounds in multiple cell lines (Supplementary information, Table S1). GC-376
(64), boceprevir (28), and calpain inhibitor II (61) were well tolerated and had CC50 values of over 100 µM for all the cell lines tested. MG-132 (43) was cytotoxic to all the cells with
CC50 values less than 1 µM except A549 cells. Calpain inhibitor XII (62) had acceptable cellular cytotoxicity with CC50 values above 50 µM for all the cell lines tested (Supplementary
information, Table S1). Next, we chose four compounds boceprevir (28), calpain inhibitors II (61), XII (62), and GC-376 (64) for the antiviral assay with infectious SARS-CoV-2. MG-132 (43)
was not included due to its cytotoxicity. Gratifyingly, all four compounds showed potent antiviral activity against SARS-CoV-2 in the primary viral cytopathic effect (CPE) assay with 50%
effective concentration (EC50) values ranging from 0.49 to 3.37 µM (Fig. 5a–d and Table 3). Their antiviral activity was further confirmed in the secondary viral yield reduction (VYR) assay
(Fig. 5e–h and Table 3). The most potent compound was calpain inhibitor XII (62), which showed an EC50 of 0.49 µM in the primary CPE assay and an EC50 of 0.78 µM in the secondary VYR assay
(Table 3; Supplementary information, Fig. S2). In comparison, remdesivir was reported to inhibit SARS-CoV-2 in the VYR assay with an EC50 of 0.77 µM.20 None of the compounds inhibited the
unrelated influenza virus A/California/07/2009 (H1N1) virus (EC50 > 20 µM) (Supplementary information, Table S1), suggesting that the antiviral activity of the four compounds (boceprevir,
calpain inhibitors II, XII, and GC-376) against SARS-CoV-2 is specific. In comparison with recently reported SARS-CoV-2 Mpro inhibitors (Table 3), the hits identified herein represent some
of the most potent and selective drug candidates with broad chemical diversity. COMPLEX CRYSTAL STRUCTURE OF SARS-COV-2 MPRO WITH GC-376 (64) The crystal structure of the SARS-CoV-2 Mpro in
complex with GC-376 (64) was solved in the P3221 space group at 2.15 Å resolution (PDB: 6WTT) (Supplementary information, Table S2). There are three protomers per asymmetric unit (ASU) with
two constituting a biological dimer and the third forming a dimer with a crystallographic symmetry related neighboring protomers (Supplementary information, Fig. S3). The presence of three
protomers in our crystal structure allowed us to capture different binding configurations of GC-376 (64) (Fig. 6), a unique feature that was not observed in previous X-ray crystal
structures.8,10,11 The pairwise r.m.s.d among the protomer backbone Cα atoms ranges from 0.435 to 0.564 Å. Previously, SARS-CoV Mpro and SARS-CoV-2 Mpro crystal structures have been solved
most frequently as one protomer per ASU, and occasionally a dimer.8,10,11,21,22 In its native state, Mpro requires dimerization to become catalytically active23,24, which is supported by our
native MS data (Supplementary information, Fig. S1). In our crystal structures, all three protomers appear catalytically competent, with the third protomer activated by the N-finger from an
adjacent ASU. The well-defined electron density also clearly shows a serine at the protease N-terminus, again indicating that the first methionine in the protease construct was cleaved
(Supplementary information, Fig. S3b). GC-376 (64) forms an extensive network of hydrogen bonds with the active site while also exhibiting excellent geometric complementarity (Fig. 6). These
interactions are coupled with the thermodynamic payoff of covalent adduct formation between the aldehyde bisulfite warhead and Cys145, making GC-376 (64) one of the most potent SARS-CoV-2
Mpro inhibitors in vitro with an IC50 of 0.030 ± 0.008 µM (Table 3). Along with other known Mpro inhibitors (N3, 13B, 11A and 11B) (Table 3), GC-376 (64) mimics the peptide substrate that is
cleaved by this enzyme (Fig. 6e).8,22,25,26 The glutamine surrogate γ-lactam ring is a cyclized derivative of the P1 glutamine side chain that normally occupies the S1 site; here it forms
hydrogen bonds with the His163 and Glu166 side chains and the main chain of Phe140 (Fig. 6a, c). An amide bond connects the γ-lactam side chain to an isobutyl moiety that embeds itself in
the hydrophobic S2 site formed by His41, Met49, and Met169. Normally, this S2 site in SARS-CoV-2 Mpro can accommodate a variety of hydrophobic substitutions such as isobutyl in GC-376 (64)
and N3), cyclopropyl in (13B), cyclohexyl in (11A), and 3-fluorophenyl in (11B) (Table 3).27,28 A carbamate bond in GC-376 (64), which forms hydrogen bonds with the main chain of Glu166 and
the side chain of Gln189, connects the P2 isobutyl group to a phenylmethyl ester that interacts with the aliphatic S4 site. Compared with previous inhibitors, the phenylmethyl ester of
GC-376 (64) exhibits high complementarity with the S4 site, and the extensive non-polar interactions may contribute significantly to the potency of this compound (Fig. 6e). Three copies of
GC-376 (64) were found in the crystal structure, one in each protomer active site (Fig. 6a, c; Supplementary information, Fig. S4). The configurations of GC-376 (64) were consistent in
protomers A and B, where the thioacetal hydroxide is positioned in the “oxyanion hole” formed by the backbone amides of Gly143, Ser144, and Cys145 (Fig. 6a; Supplementary information, Fig.
S4), resulting in the (S)-configuration. It is noted that aldehydes (11A and 11B) also bind the active site of SARS-CoV-2 Mpro in the (S)-configuration (PDB: 6M0K and 6LZE) (Supplementary
information, Fig. S5a).11 In protomer C, however, the same hydroxide group orients outwards from the oxyanion hole, forming hydrogen bonds with His41 (Fig. 6c), which gives the
(R)-configuration. This (R)-configuration is consistent with the binding mode of α-ketoamide (13B) in the active site of SARS-CoV-2 Mpro (PDB: 6Y2F) (Supplementary information, Fig. S5b).8
These two unique configurations R and S might be a result of the Cys145 thiol nucleophilic attacking the aldehyde of GC-376 (64) from two different faces (Fig. 6b, d). The fact that GC-376
(64) can adopt two different configurations R and S upon binding to the active site might explain its high binding affinity towards the target. An additional difference between the
configurations of GC-376 (64) in protomers A, B and C is observed in the orientation of the phenylmethyl ester. In protomer C, the CH2 of the phenylmethyl points toward the main chain of
Leu167 in a “_cis_” conformation (Fig. 6c), whereas in protomers A and B this same CH2 points downwards in a “_trans_” conformation (Fig. 6a; Supplementary information, Fig. S4).
Consequently, this influences the rotameric configuration of the Leu167 isobutyl moiety, where a rotational adjustment of 180° occurs at its “β” carbon. Furthermore, large rearrangements are
observed in the flexible loop that consists of residues 45–51 (TSEDMLN in SARS-CoV-2 Mpro) and forms the S2 and S3′ subsites (Fig. 6f), explaining the broad substrate scope in the P2 site
(Table 3). The loop conformations in protomers B and C may be influenced by crystal packing interactions with protomers from adjacent ASUs and resemble the conformations in previously
determined structures.10,11 Meanwhile, the conformation of protomer A is less restrained and exhibits the most significant conformational divergence. The different loop conformations offer a
glimpse of the protein plasticity that allows Mpro to accommodate peptides with differing amino acid composition, and underscores the importance of considering this flexibility when
analyzing and modeling protein-ligand interactions for Mpro. DISCUSSION Coronaviruses have caused three epidemics/pandemics in the past 20 years including SARS, MERS, and COVID-19. With the
ongoing pandemic of COVID-19, scientists and researchers around the globe are racing to find effective vaccines and antiviral drugs.6 The viral polymerase inhibitor remdesivir holds the
greatest promise and it is currently being evaluated in several clinical trials.29,30 Remdesivir was recently granted emergency use authorization by FDA. The HIV drug combination lopinavir
and ritonavir recently failed in a clinical trial for COVID-19 as no significant therapeutic efficacy was observed.31 To address this unmet medical need, we initiated a drug repurposing
screening to identify potent inhibitors against the SARS-CoV-2 Mpro from a collection of FDA-approved protease inhibitors. The Mpro has been shown to be a validated antiviral drug target for
SARS-CoV and MERS-CoV.32 As the SARS-CoV-2 Mpro shares a high sequence similarity with SARS-CoV and to a less extent with MERS-CoV, we reasoned that inhibiting the enzymatic activity of
SARS-CoV-2 Mpro will similarly prevent viral replication.8,10 Noticeable findings from our study include: (1) Boceprevir (28), an FDA-approved HCV drug, inhibits the enzymatic activity of
Mpro with IC50 of 4.13 µM, and has an EC50 of 1.90 µM against the SARS-CoV-2 virus in the cellular viral CPE assay. The therapeutic potential of boceprevir (28) should be further evaluated
in relevant animal models and human clinical trials. Since boceprevir (28) is an FDA-approved drug, the dose, toxicity, formulation and pharmacokinetic properties are already known, which
will greatly speed up the design of follow-up studies; (2) GC-376 (64), an investigational veterinary drug, showed promising antiviral activity against the SARS-CoV-2 virus (EC50 = 3.37 µM).
It has the highest enzymatic inhibition against the Mpro with an IC50 value of 0.03 µM. This compound has promising in vivo efficacy in treating cats infected with FIP, and has favorable in
vivo pharmacokinetic properties. Therefore, GC-376 (64) is ready to be tested in relevant animal models of SARS-CoV-2 when available. Importantly, the X-ray crystal structure of SARS-CoV-2
Mpro in complex with GC-376 (64) provides a molecular explanation of the high binding affinity of aldehyde-containing compounds as they can adopt two configurations R and S. The
conformational flexibility at the TSEDMLN loop explains the broad substrate scope at the P2 position of Mpro inhibitors; (3) Three calpain/cathepsin inhibitors, MG-132 (43), calpain
inhibitors II (61) and XII (62), are potent inhibitors of Mpro with single-digit to submicromolar efficacy in the enzymatic assay. Calpain inhibitors II (61) and XII (62) also inhibit
SARS-CoV-2 in the CPE assay with EC50 values of 2.07 and 0.49 µM, respectively. This result suggests that calpain/cathepsin inhibitors are rich sources of drug candidates for SARS-CoV-2.
Indeed, previous studies have shown that calpain II and cathepsin L are required for the proteolytic processing of the coronavirus S protein, a step that is essential for the viral fusion
and genome release during the early stage of viral replication.33 Calpain and cathepsin inhibitors such as MDL28170 (calpain inhibitor III),33 MG-132,34 calpain inhibitor VI35 have been
shown to inhibit SARS-CoV replication in cell culture. Other than the increased potency of targeting both Mpro and calpain/cathepsin, an additional benefit of such dual inhibitors might be
their high genetic barrier to drug resistance. A significant number of calpain/cathepsin inhibitors have been developed over the years for various diseases including cancer,
neurodegeneration disease, kidney diseases, and ischemia/reperfusion injury.36 Given our promising results of calpain inhibitors II (61) and XII (62) in inhibiting the SARS-CoV-2 Mpro and
their potent antiviral activity in cell culture, it might be worthwhile to repurpose them as antivirals for SARS-CoV-2. All potent SARS-CoV-2 Mpro inhibitors contain reactive warheads such
as α-ketoamide (boceprevir (28), calpain inhibitor XII (62)) or aldehyde (MG-132 (43), calpain inhibitor II (61)) or aldehyde prodrug, the bisulfite (GC-376 (64)). This result suggests that
reactive warheads might be essential for SARS-CoV-2 Mpro inhibition. The compounds identified in this study represent some of the most potent and selective hits reported so far, and have
similar activity to the recently reported SARS-CoV-2 Mpro inhibitors (EBSELEN, N3, and 13B) (Table 3). Calpain inhibitor XII (62) had similar potency to the recently disclosed compounds (11A
and 11B) (Table 3).11 Notably, boceprevir (28), calpain inhibitor II (61) and XII (62) have different chemical scaffolds than the substrate mimetics GC-376 (64), (N3, 13B, 11A, and 11B),
therefore providing new opportunities for designing more potent and selective SARS-CoV-2 Mpro inhibitors. Aside from the above positive results, we also showed that ritonavir (9) and
lopinavir (10) failed to inhibit the SARS-CoV-2 Mpro (IC50 > 20 µM, Fig. 2), which might explain why they lack efficacy in clinical trials for COVID-19.31 Camostat (39) was recently
proposed to inhibit SARS-CoV-2 entry through inhibiting the host TMPRSS2, a host serine protease that is important for viral S protein priming.37 In our study, we found that camostat (39)
has no inhibition against the SARS-CoV-2 Mpro (IC50 > 20 µM). In summary, this study identified several potent SARS-CoV-2 Mpro inhibitors with potent enzymatic inhibition as well as
cellular antiviral activity. Further development based on these hits might lead to clinically useful COVID-19 antivirals. They can be used either alone or in combination with polymerase
inhibitors such as remdesivir as a means to achieve potential synergistic antiviral effect as well as to suppress drug resistance. MATERIALS AND METHODS CELL LINES AND VIRUSES Human
rhabdomyosarcoma RD, A549, MDCK, Caco-2, and Vero cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM); BEAS2B and HCT-8 cells were maintained in RPMI 1640 medium. Both medium
was supplemented with 10% fetal bovine serum (FBS) and 1% penicillin–streptomycin antibiotics. Cells were kept at 37 °C in a 5% CO2 atmosphere. The USA-WA1/2020 strain of SARS-CoV-2 was
obtained from the World Reference Center for Emerging Viruses and Arboviruses (WRCEVA). PROTEIN EXPRESSION AND PURIFICATION SARS-CoV-2 main protease (Mpro or 3CL) gene from strain
BetaCoV/Wuhan/WIV04/2019 was ordered from GenScript (Piscataway, NJ, USA) in the pET29a(+) vector with _E. coli_ codon optimization. pET29a(+) plasmids with SARS-CoV-2 Mpro was transformed
into competent _E. coli_ BL21(DE3) cells, and a single colony was picked to inoculate 10 mL of LB supplemented with 50 g/mL kanamycin and grown at 37 °C with shaking at 250 rpm. The 10-mL
inoculum was added to 1 L of LB with 50 g/mL kanamycin and grown to an optical density at 600 nm of 0.8, and then induced using 1.0 mM IPTG. Induced cultures were incubated at 37 °C for an
additional 3 h and then harvested, resuspended in lysis buffer (25 mM Tris, pH 7.5, 750 mM NaCl, 2 mM DTT with 0.5 mg/mL lysozyme, 0.5 mM phenylmethylsulfonyl fluoride (PMSF), 0.02 mg/mL
DNase I), and lysed by alternating sonication and French press cycles. The cell debris were removed by centrifugation at 12,000× _g_ for 45 min (20% amplitude, 1 s on/1 s off). The
supernatant was incubated with Ni-NTA resin for over 2 h at 4 °C on a rotator. The Ni-NTA resin was thoroughly washed with 30 mM imidazole in wash buffer (50 mM Tris, pH 7.0, 150 mM NaCl, 2
mM DTT); and eluted with 100 mM imidazole in elution buffer (50 mM Tris, pH 7.0, 150 mM NaCl, 2 mM DTT). The imidazole was removed via dialysis or on a 10,000-molecular-weight-cutoff
centrifugal concentrator spin column. The purity of the protein was confirmed with SDS-PAGE. The protein concentration was determined via 260 nm absorbance with ε 32890. EV-A71 2Apro and
3Cpro were expressed in the pET28b(+) vector as previously described.18,38,39 The protein sequence of the SARS-CoV-2 Mpro used in the enzymatic assay and X-ray crystallization is listed
below: SGFRKMAFPS GKVEGCMVQV TCGTTTLNGL WLDDVVYCPR HVICTSEDML NPNYEDLLIR KSNHNFLVQA GNVQLRVIGH SMQNCVLKLK VDTANPKTPK YKFVRIQPGQ TFSVLACYNG SPSGVYQCAM RPNFTIKGSF LNGSCGSVGF NIDYDCVSFC
YMHHMELPTG VHAGTDLEGN FYGPFVDRQT AQAAGTDTTI TVNVLAWLYA AVINGDRWFL NRFTTTLNDF NLVAMKYNYE PLTQDHVDIL GPLSAQTGIA VLDMCASLKE LLQNGMNGRT ILGSALLEDE FTPFDVVRQC SGVTFQ LEHHHHHH. PEPTIDE SYNTHESIS
The SARS-CoV-2 Mpro FRET substrate Dabcyl-KTSAVLQ/SGFRKME (Edans) was synthesized by solid-phase synthesis through iterative cycles of coupling and deprotection using the previously
optimized procedure.40 Specifically, chemmatrix rink-amide resin was used. Typical coupling condition was 5 equiv of amino acid, 5 equiv of HATU, and 10 equiv of DIEA in DMF for 5 min at 80
°C. For deprotection, 5% piperazine plus 0.1 M HOBt were used and the mixture was heated at 80 °C for 5 min. The peptide was cleaved from the resin using 95% TFA, 2.5% Tris and 2.5% H2O, and
the crude peptide was precipitated from ether after removal of TFA. The final peptide was purified by preparative HPLC. The purify and identity of the peptide were confirmed by analytical
HPLC (> 98% purity) and mass spectrometry. [M + 3]3+ was calculated as 694.15 and detected as 694.90; [M + 4]4+ was calculated as 520.86 and detected as 521.35. EDANS STANDARD CURVE Edans
standard curve was generated as described below: 200 nM SARS-CoV-2 Mpro was incubated with varying concentrations of FRET substrate (0.5–40 µM) and the reaction progress was monitored until
the fluorescence signals reached plateau, at which point we deemed all the FRET substrate was digested by Mpro. The endpoint fluorescence signal was plotted against FRET substrate
concentration with a linear regression function in Prism 5. NATIVE MASS SPECTROMETRY Prior to analysis, the protein was buffer exchanged into 0.2 M ammonium acetate (pH 6.8) and diluted to
10 μM. DTT was dissolved in water and prepared at a 400 mM stock. Each ligand was dissolved in ethanol and diluted to 10× stock concentrations. The final mixture was prepared by adding 4 μL
protein, 0.5 μL DTT stock, and 0.5 μL ligand stock for final concentration of 4 mM DTT and 8 μM protein. Final ligand concentrations were used as annotated. The mixtures were then incubated
for 10 min at room temperature prior to analysis. Each sample was mixed and analyzed in triplicate. Native MS was performed using a Q-Exactive HF quadrupole-Orbitrap mass spectrometer with
the Ultra-High Mass Range research modifications (Thermo Fisher Scientific). Samples were ionized using nano-electrospray ionization in positive ion mode using 1.0 kV capillary voltage at a
150 °C capillary temperature. The samples were all analyzed with a 1000–25,000 m/z range, the resolution set to 30,000, and a trapping gas pressure set to 3. Source fragmentation (10–50 V)
was applied to all samples to aid in desolvation. Data were deconvolved and analyzed with UniDec.41 ENZYMATIC ASSAYS For reaction condition optimization, 200 µM SARS-CoV-2 Mpro was used. pH
6.0 buffer contains 20 mM MES, pH 6.0, 120 mM NaCl, 0.4 mM EDTA, 4 mM DTT and 20% glycerol; pH 6.5 buffer contains 20 mM HEPES, pH 6.5, 120 mM NaCl, 0.4 mM EDTA, 4 mM DTT and 20% glycerol;
pH 7.0 buffer contains 20 mM HEPES, pH 7.0, 120 mM NaCl, 0.4 mM EDTA, 4 mM DTT and 20% glycerol. Upon addition of 20 µM FRET substrate, the reaction progress was monitored for 1 h. The first
15 min of reaction was used to calculate initial velocity via linear regression in Prism 5. Mpro displays the highest proteolytic activity in pH 6.5 buffer. All the following enzymatic
assays were carried out in pH 6.5 buffer. For the measurements of _K_m/_V_max, screening of the protease inhibitor library, as well as IC50 measurements, proteolytic reaction with 100 nM
Mpro in 100 µL of pH 6.5 reaction buffer was carried out at 30 °C in a Cytation 5 imaging reader (Thermo Fisher Scientific) with filters for excitation at 360/40 nm and emission at 460/40
nm. Reactions were monitored every 90 s. For _K_m/_V_max measurements, a FRET substrate concentration ranging from 0 to 200 µM was applied. The initial velocity of the proteolytic activity
was calculated by linear regression for the first 15 min of the kinetic progress curves. The initial velocity was plotted against the FRET concentration with the classic Michaelis–Menten
equation in Prism 5 software. For the screening of protease inhibitor library and IC50 measurements, 100 nM Mpro was incubated with protease inhibitor at 30 °C for 30 min in reaction buffer,
then the reaction was initiated by adding 10 µM FRET substrate, the reaction was monitored for 1 h, and the initial velocity was calculated for the first 15 min by linear regression. The
IC50 was calculated by plotting the initial velocity against various concentrations of protease inhibitors by use of a dose-response curve in Prism 5 software. Proteolytic reaction progress
curve kinetics measurements with GC376, MG132, boceprevir, calpain inhibitor II, and calpain inhibitor XII used for curve fitting, were carried out as follows: 5 nM Mpro protein was added to
20 µM FRET substrate with various concentrations of testing inhibitor in 200 µL of reaction buffer at 30 °C to initiate the proteolytic reaction. The reaction was monitored for 4 h. The
progress curves were fit to a slow binding Morrison equation (Eq. (3)) as described previously18,42: $$E + I \,\,{\mathop {\leftrightarrow}\limits _{k_1} ^{K_2}}\,\, EI \,\,{\mathop
{\leftrightarrow}\limits _{k_1} ^{K_2}}\,\, EI \,\,^{\ast}$$ (1) $$\mathrm{K}_{\mathrm{I}} = \mathrm{k}_{ - 1}/\mathrm{k}_1$$ (2) $$\mathrm{P}\left( \mathrm{t} \right) = \mathrm{P}_0 +
\mathrm{V}_{\mathrm{s}}{\mathrm{t}}-\left( {\mathrm{Vs}-\mathrm{V}_0} \right)\left( {1-e^{ - \mathrm{kt}}} \right)/\mathrm{k}$$ (3) $$\mathrm{k} = \mathrm{k}_2\left[ \mathrm{I}
\right]/\left( {\mathrm{K}_{\mathrm{I}} + \left[ \mathrm{I} \right]} \right)$$ (4) where P(t) is the fluorescence signal at time t, P0 is the background signal at time 0, V0, Vs, and k
represent, respectively, the initial velocity, the final steady-state velocity and the apparent first-order rate constant for the establishment of the equilibrium between EI and EI*.42 k2/KI
is commonly used to evaluate the efficacy for covalent inhibitor. We observed substrate depletion when proteolytic reactions progress longer than 90 min; therefore only first 90 min of the
progress curves were used in the curve fitting (Fig. 6, middle column). In this study, we could not accurately determine the k2 for the protease inhibitors: calpain inhibitor II, MG132,
boceprevir, and calpain inhibitor XII, due to the very slow k2 in these cases: significant substrate depletion before the establishment of the equilibrium between EI and EI*. In these cases,
KI was determined with Morrison equation in Prism 5. DIFFERENTIAL SCANNING FLUORIMETRY The binding of protease inhibitors to Mpro was monitored by differential scanning fluorimetry (DSF)
using a Thermo Fisher QuantStudioTM 5 Real-Time PCR System. TSA plates were prepared by mixing Mpro protein (final concentration of 3 μM) with inhibitors, and incubated at 30 °C for 30 min.
1× SYPRO orange (Thermo Fisher Scientific) were added and the fluorescence of the plates were taken under a temperature gradient ranging from 20 to 90 °C (incremental steps of 0.05 °C/s).
The melting temperature (Tm) was calculated as the mid-log of the transition phase from the native to the denatured protein using a Boltzmann model (Protein Thermal Shift Software v1.3).
Thermal shift that was represented as ΔTm was calculated by subtracting reference melting temperature of proteins in DMSO from the Tm in the presence of compound. CYTOTOXICITY MEASUREMENT
A549, MDCK, HCT-8, Caco-2, Vero, and BEAS2B cells for cytotoxicity CPE assays were seeded and grown overnight at 37 °C in a 5% CO2 atmosphere to ∼90% confluence on the next day. Cells were
washed with phosphate-buffered saline (PBS) and 200 µL DMEM containing 2% FBS and 1% penicillin–streptomycin, and various concentrations of protease inhibitors were added to each well. 48 h
after addition of the protease inhibitors, cells were stained with 66 μg/mL neutral red for 2 h, and neutral red uptake was measured at an absorbance at 540 nm using a Multiskan FC
microplate photometer (Thermo Fisher Scientific). The CC50 values were calculated from best-fit dose-response curves using GraphPad Prism 5 software. SARS-COV-2 CPE ASSAY Antiviral
activities of test compounds were determined in nearly confluent cultures of Vero 76 cells. The assays were performed in 96-well Corning microplates. Cells were infected with approximately
60-fold of 50% cell culture infectious doses (CCID50) of SARS-CoV-2 and 50% effective concentrations (EC50) were calculated based on virus-induced CPE quantified by neutral red dye uptake
after 5 days of incubation. USA-WA1/2020 strain of SARS-CoV-2 obtained from the WRCEVA was used for both CPE and VYR assays. Three microwells at each concentration of compound were infected.
Two uninfected microwells served as toxicity controls. Cells were stained for viability with neutral red (0.11% final concentration) for 2 h. Excess dye was rinsed from the cells with PBS.
The absorbed dye was eluted from the cells with 0.1 mL of 50% Sörensen’s citrate buffer (pH 4.2)-50% ethanol. Plates were read for optical density determination at 540 nm. Readings were
converted to the percentage of the results for the uninfected control using an Excel spreadsheet developed for this purpose. EC50 values were determined by plotting percent CPE versus log10
inhibitor concentration. Toxicity at each concentration was determined in uninfected wells in the same microplates by measuring dye uptake. SARS-COV-2 VYR ASSAY VYR assays were conducted by
first replicating the viruses in the presence of test compound. Supernatant was harvested 3 days post infection from each concentration of test compound and the virus yield was determined by
endpoint dilution method. Briefly, supernatant virus was serially diluted in log10 increments and then plated onto quadruplicate wells of 96-well plates seeded with Vero 76 cells. The
presence or absence of CPE for determining a viral endpoint was evaluated by microscopic examination of cells 6 days post infection. From these data, 90% virus inhibitory concentrations
(EC90) were determined by regression analysis. INFLUENZA A VIRUS A/CALIFORNIA/07/2009 (H1N1) PLAQUE REDUCTION ASSAY The plaque assay was performed according to previously published
procedures.43 MPRO CRYSTALLIZATION AND STRUCTURE DETERMINATION 10 mg/mL of SARS-CoV-2 Mpro was incubated with 2 mM GC376 at 4 °C overnight. The protein was diluted to 2.5 mg/mL the following
day in protein buffer (50 mM Tris, pH 7.0, 150 mM NaCl, 4 mM DTT). Since GC3760 is water soluble, no precipitation was observed, and centrifugation was not necessary. Crystals were grown by
mixing 2 μL of the protein solution with 1 μL of the precipitant solution (15% PEG 2 K, 10% 1,6-hexanediol, and 0.2 M NaCl) in a hanging-drop vapor-diffusion apparatus. Crystals were
cryoprotected by transferring to a cryoprotectant solution (20% PEG 2 K, 10% 1,6-hexanediol, 20% glycerol) and flash-frozen in liquid nitrogen. X-ray diffraction data for the SARS-CoV-2
Mpro–GC376 complex structure was collected on the SBC 19-ID beamline at the Advanced Photon Source (APS) in Argonne, IL, and processed with the HKL3000 software suite.44 The CCP4 versions of
MOLREP was used for molecular replacement using a previously solved SARS-CoV-2 Mpro structure (PDB ID: 5RGG) as a reference model.45 Rigid and restrained refinements were performed using
REFMAC and model building was performed with COOT.46,47 Protein structure figures were made using PyMOL (Schrödinger, LLC). DATA AVAILABILITY The structure for SARS-CoV-2 Mpro has been
deposited in the Protein Data Bank with accession number 6WTT. REFERENCES * Mahase, E. Coronavirus: covid-19 has killed more people than SARS and MERS combined, despite lower case fatality
rate. _BMJ_ 368, m641 (2020). PubMed Google Scholar * Lu, R. et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor
binding. _Lancet_ 395, 565–574 (2020). CAS PubMed PubMed Central Google Scholar * Wu, A. et al. Genome composition and divergence of the novel coronavirus (2019-nCoV) originating in
China. _Cell Host Microbe_ 27, 325–328 (2020). CAS PubMed PubMed Central Google Scholar * Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species
Severe acute respiratory syndrome-related coronavirus: classifying 2019-nCoV and naming it SARS-CoV-2. _Nat. Microbiol_. 5, 536–544 (2020). * Tang, B. et al. An updated estimation of the
risk of transmission of the novel coronavirus (2019-nCov). _Infect. Dis. Model._ 5, 248–255 (2020). PubMed PubMed Central Google Scholar * Li, G. & De Clercq, E. Therapeutic options
for the 2019 novel coronavirus (2019-nCoV). _Nat. Rev. Drug Discov._ 19, 149–150 (2020). PubMed Google Scholar * Xia, S. et al. Inhibition of SARS-CoV-2 (previously 2019-nCoV) infection by
a highly potent pan-coronavirus fusion inhibitor targeting its spike protein that harbors a high capacity to mediate membrane fusion. _Cell Res._ 30, 343–355 (2020). CAS PubMed PubMed
Central Google Scholar * Zhang, L. et al. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved alpha-ketoamide inhibitors. _Science_ 368, 409–412 (2020).
CAS PubMed PubMed Central Google Scholar * Anand, K., Ziebuhr, J., Wadhwani, P., Mesters, J. R. & Hilgenfeld, R. Coronavirus main proteinase (3CLpro) structure: basis for design of
anti-SARS drugs. _Science_ 300, 1763–1767 (2003). CAS PubMed Google Scholar * Jin, Z. et al. Structure of M(pro) from SARS-CoV-2 and discovery of its inhibitors. _Nature_ 582, 289–293
(2020). CAS PubMed Google Scholar * Dai, W. et al. Structure-based design of antiviral drug candidates targeting the SARS-CoV-2 main protease. _Science_
https://doi.org/10.1126/science.abb4489 (2020). Article PubMed PubMed Central Google Scholar * Wingfield, P. T. N-terminal methionine processing. _Curr. Protoc. Protein Sci._ 88,
6.14.1–6.14.3 (2017). CAS Google Scholar * Pedersen, N. C. et al. Efficacy of a 3C-like protease inhibitor in treating various forms of acquired feline infectious peritonitis. _J. Feline
Med. Surg._ 20, 378–392 (2018). PubMed PubMed Central Google Scholar * Kim, Y. et al. Reversal of the progression of fatal coronavirus infection in cats by a broad-spectrum coronavirus
protease inhibitor. _PLoS Pathog._ 12, e1005531 (2016). PubMed PubMed Central Google Scholar * Kim, Y. et al. Broad-spectrum antivirals against 3C or 3C-like proteases of picornaviruses,
noroviruses, and coronaviruses. _J. Virol._ 86, 11754–11762 (2012). CAS PubMed PubMed Central Google Scholar * Hayden, F. G. et al. Phase II, randomized, double-blind, placebo-controlled
studies of ruprintrivir nasal spray 2-percent suspension for prevention and treatment of experimentally induced rhinovirus colds in healthy volunteers. _Antimicrob. Agents Chemother._ 47,
3907–3916 (2003). CAS PubMed PubMed Central Google Scholar * Shie, J. J. et al. Inhibition of the severe acute respiratory syndrome 3CL protease by peptidomimetic alpha,beta-unsaturated
esters. _Bioorg. Med. Chem._ 13, 5240–5252 (2005). CAS PubMed PubMed Central Google Scholar * Musharrafieh, R. et al. Validating enterovirus D68-2A(pro) as an antiviral drug target and
the discovery of telaprevir as a potent D68-2A(pro) inhibitor. _J. Virol._ 93, e0221-18 (2019). Google Scholar * Kuo, C. J. et al. Design, synthesis, and evaluation of 3C protease
inhibitors as anti-enterovirus 71 agents. _Bioorg. Med. Chem._ 16, 7388–7398 (2008). CAS PubMed PubMed Central Google Scholar * Wang, M. et al. Remdesivir and chloroquine effectively
inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. _Cell Res._ 30, 269–271 (2020). CAS PubMed PubMed Central Google Scholar * Yang, H. et al. The crystal structures of
severe acute respiratory syndrome virus main protease and its complex with an inhibitor. _Proc. Natl. Acad. Sci. USA_ 100, 13190–13195 (2003). CAS PubMed PubMed Central Google Scholar *
Muramatsu, T. et al. SARS-CoV 3CL protease cleaves its C-terminal autoprocessing site by novel subsite cooperativity. _Proc. Natl Acad. Sci. USA_ 113, 12997–13002 (2016). CAS PubMed PubMed
Central Google Scholar * Li, C. et al. Maturation mechanism of severe acute respiratory syndrome (SARS) coronavirus 3C-like proteinase. _J. Biol. Chem._ 285, 28134–28140 (2010). CAS
PubMed PubMed Central Google Scholar * Anand, K. et al. Structure of coronavirus main proteinase reveals combination of a chymotrypsin fold with an extra alpha-helical domain. _EMBO J._
21, 3213–3224 (2002). CAS PubMed PubMed Central Google Scholar * Xue, X. et al. Structures of two coronavirus main proteases: implications for substrate binding and antiviral drug
design. _J. Virol._ 82, 2515–2527 (2008). CAS PubMed Google Scholar * Zhang, L. et al. Alpha-ketoamides as broad-spectrum inhibitors of coronavirus and enterovirus replication:
structure-based design, synthesis, and activity assessment. _J. Med. Chem._ 63, 4562–4578 (2020). CAS PubMed Google Scholar * Chuck, C. P. et al. Profiling of substrate specificity of
SARS-CoV 3CL. _PLoS ONE_ 5, e13197 (2010). PubMed PubMed Central Google Scholar * Hegyi, A. & Ziebuhr, J. Conservation of substrate specificities among coronavirus main proteases. _J.
Gen. Virol._ 83, 595–599 (2002). PubMed Google Scholar * Grein, J. et al. Compassionate use of remdesivir for patients with severe Covid-19. _N. Engl. J. Med._ 382, 2327–2336 (2020). CAS
PubMed Google Scholar * Holshue, M. L. et al. First case of 2019 novel coronavirus in the United States. _N. Engl. J. Med._ 382, 929–936 (2020). CAS PubMed PubMed Central Google
Scholar * Cao, B. et al. A trial of lopinavir–ritonavir in adults hospitalized with severe Covid-19. _N. Engl. J. Med._ 382, 1787–1799 (2020). PubMed Google Scholar * Zumla, A., Chan, J.
F., Azhar, E. I., Hui, D. S. & Yuen, K. Y. Coronaviruses—drug discovery and therapeutic options. _Nat. Rev. Drug Discov._ 15, 327–347 (2016). CAS PubMed PubMed Central Google Scholar
* Simmons, G. et al. Inhibitors of cathepsin L prevent severe acute respiratory syndrome coronavirus entry. _Proc. Natl Acad. Sci. USA_ 102, 11876–11881 (2005). CAS PubMed PubMed Central
Google Scholar * Schneider, M. et al. Severe acute respiratory syndrome coronavirus replication is severely impaired by MG132 due to proteasome-independent inhibition of M-calpain. _J.
Virol._ 86, 10112–10122 (2012). CAS PubMed PubMed Central Google Scholar * Barnard, D. L. et al. Inhibition of severe acute respiratory syndrome-associated coronavirus (SARSCoV) by
calpain inhibitors and beta-D-N4-hydroxycytidine. _Antivir. Chem. Chemother._ 15, 15–22 (2004). CAS PubMed Google Scholar * Ono, Y., Saido, T. C. & Sorimachi, H. Calpain research for
drug discovery: challenges and potential. _Nat. Rev. Drug Discov._ 15, 854–876 (2016). CAS PubMed Google Scholar * Hoffmann, M. et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2
and is blocked by a clinically proven protease inhibitor. _Cell_ 181, 271–280 (2020). CAS PubMed PubMed Central Google Scholar * Shang, L. et al. Biochemical characterization of
recombinant Enterovirus 71 3C protease with fluorogenic model peptide substrates and development of a biochemical assay. _Antimicrob. Agents Chemother._ 59, 1827–1836 (2015). CAS PubMed
PubMed Central Google Scholar * Cai, Q. et al. Conformational plasticity of the 2A proteinase from enterovirus 71. _J. Virol._ 87, 7348–7356 (2013). CAS PubMed PubMed Central Google
Scholar * Cady, S. D., Wang, J., Wu, Y., DeGrado, W. F. & Hong, M. Specific binding of adamantane drugs and direction of their polar amines in the pore of the influenza M2 transmembrane
domain in lipid bilayers and dodecylphosphocholine micelles determined by NMR spectroscopy. _J. Am. Chem. Soc._ 133, 4274–4284 (2011). CAS PubMed PubMed Central Google Scholar * Marty,
M. T. et al. Bayesian deconvolution of mass and ion mobility spectra: from binary interactions to polydisperse ensembles. _Anal. Chem._ 87, 4370–4376 (2015). CAS PubMed PubMed Central
Google Scholar * Morrison, J. F. & Walsh, C. T. The behavior and significance of slow-binding enzyme inhibitors. _Adv. Enzymol. Relat. Areas Mol. Biol._ 61, 201–301 (1988). CAS PubMed
Google Scholar * Ma, C., Zhang, J. & Wang, J. Pharmacological characterization of the spectrum of antiviral activity and genetic barrier to drug resistance of M2-S31N channel
blockers. _Mol. Pharmacol._ 90, 188–198 (2016). CAS PubMed PubMed Central Google Scholar * Minor, W., Cymborowski, M., Otwinowski, Z. & Chruszcz, M. HKL-3000: the integration of data
reduction and structure solution—from diffraction images to an initial model in minutes. _Acta Crystallogr. D Biol. Crystallogr._ 62, 859–866 (2006). PubMed Google Scholar * Vagin, A.
& Teplyakov, A. Molecular replacement with MOLREP. _Acta Crystallogr. D Biol. Crystallogr._ 66, 22–25 (2010). CAS PubMed Google Scholar * Murshudov, G. N. et al. REFMAC5 for the
refinement of macromolecular crystal structures. _Acta Crystallogr. D Biol. Crystallogr._ 67, 355–367 (2011). CAS PubMed PubMed Central Google Scholar * Emsley, P. & Cowtan, K. Coot:
model-building tools for molecular graphics. _Acta Crystallogr. D Biol. Crystallogr._ 60, 2126–2132 (2004). PubMed Google Scholar * Bonuccelli, G. et al. Proteasome inhibitor (MG-132)
treatment of mdx mice rescues the expression and membrane localization of dystrophin and dystrophin-associated proteins. _Am. J. Pathol._ 163, 1663–1675 (2003). CAS PubMed PubMed Central
Google Scholar * Mani, S. K. et al. In vivo administration of calpeptin attenuates calpain activation and cardiomyocyte loss in pressure-overloaded feline myocardium. _Am. J. Physiol. Heart
Circ. Physiol._ 295, H314–326 (2008). CAS PubMed PubMed Central Google Scholar * Peng, S., Kuang, Z., Zhang, Y., Xu, H. & Cheng, Q. The protective effects and potential mechanism of
Calpain inhibitor Calpeptin against focal cerebral ischemia-reperfusion injury in rats. _Mol. Biol. Rep._ 38, 905–912 (2011). CAS PubMed Google Scholar * Akdemir, O. et al. Therapeutic
efficacy of SJA6017, a calpain inhibitor, in rat spinal cord injury. _J. Clin. Neurosci._ 15, 1130–1136 (2008). CAS PubMed Google Scholar * Li, S. Z. et al. ALLN hinders HCT116 tumor
growth through Bax-dependent apoptosis. _Biochem. Biophys. Res. Commun._ 437, 325–330 (2013). CAS PubMed Google Scholar * Delli Pizzi, S. et al. Morphological and metabolic changes in the
nigro-striatal pathway of synthetic proteasome inhibitor (PSI)-treated rats: a MRI and MRS study. _PLoS ONE_ 8, e56501 (2013). CAS PubMed PubMed Central Google Scholar Download
references ACKNOWLEDGEMENTS This research was partially supported by the National Institutes of Health (NIH) (Grant AI147325) and the Arizona Biomedical Research Centre Young Investigator
grant (ADHS18-198859) to J.W., B.H. and B.T. thanks for the support from the Respiratory Diseases Branch, National Institute of Allergy and Infectious Diseases, NIH, USA (Contract
N01-AI-30048). J.A.T. and M.T.M. were funded by the National Institute of General Medical Sciences, NIH (Grant R35 GM128624 to M.T.M.). We thank Michael Kemp for assistance with
crystallization and X-ray diffraction data collection. We also thank the staff members of the Advanced Photon Source of Argonne National Laboratory, particularly those at the Structural
Biology Center (SBC), with X-ray diffraction data collection. SBC-CAT is operated by UChicago Argonne, LLC, for the U.S. Department of Energy, Office of Biological and Environmental Research
under contract DE-AC02-06CH11357. AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Department of Pharmacology and Toxicology, College of Pharmacy, The University of Arizona, Tucson, AZ, 85721,
USA Chunlong Ma, Yanmei Hu, Tommy Szeto & Jun Wang * Department of Molecular Medicine, Morsani College of Medicine, University of South Florida, Tampa, FL, 33612, USA Michael Dominic
Sacco, Xiujun Zhang & Yu Chen * Institute for Antiviral Research, Utah State University, Logan, UT, 84322, USA Brett Hurst & Bart Tarbet * Department of Animal, Dairy and Veterinary
Sciences, Utah State University, Logan, UT, 84322, USA Brett Hurst & Bart Tarbet * Department of Chemistry and Biochemistry, The University of Arizona, Tucson, AZ, 85721, USA Julia Alma
Townsend & Michael Thomas Marty Authors * Chunlong Ma View author publications You can also search for this author inPubMed Google Scholar * Michael Dominic Sacco View author
publications You can also search for this author inPubMed Google Scholar * Brett Hurst View author publications You can also search for this author inPubMed Google Scholar * Julia Alma
Townsend View author publications You can also search for this author inPubMed Google Scholar * Yanmei Hu View author publications You can also search for this author inPubMed Google Scholar
* Tommy Szeto View author publications You can also search for this author inPubMed Google Scholar * Xiujun Zhang View author publications You can also search for this author inPubMed
Google Scholar * Bart Tarbet View author publications You can also search for this author inPubMed Google Scholar * Michael Thomas Marty View author publications You can also search for this
author inPubMed Google Scholar * Yu Chen View author publications You can also search for this author inPubMed Google Scholar * Jun Wang View author publications You can also search for
this author inPubMed Google Scholar CONTRIBUTIONS J.W. and C.M. conceived and designed the study; C.M. expressed the Mpro with the assistance of T.S.; C.M. performed the primary screening,
secondary IC50 determination, thermal shift binding assay, and enzymatic kinetic studies; M.D.S. carried out Mpro crystallization and structure determination with the assistance of X.Z., and
analyzed the data with Y.C.; B.H. and B.T. performed the SARS-CoV-2 CPE and VYR assays; J.A.T. performed the native mass spectrometry experiments with the guidance from M.T.M.; Y.H.
performed the plaque reduction assay with influenza A/California/07/2009 (H1N1) virus; J.W. and Y.C. secured funding and supervised the study; J.W., Y.C., and C.M. wrote the paper with the
input from others. CORRESPONDING AUTHORS Correspondence to Yu Chen or Jun Wang. ETHICS DECLARATIONS COMPETING INTERESTS J.W. and C.M. are inventors of a pending patent that claims the use of
the identified compounds for COVID-19. SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION FIGURE S1 SUPPLEMENTARY INFORMATION FIGURE S2 SUPPLEMENTARY INFORMATION FIGURE S3 SUPPLEMENTARY
INFORMATION FIGURE S4 SUPPLEMENTARY INFORMATION FIGURE S5 SUPPLEMENTARY INFORMATION, TABLE S1 SUPPLEMENTARY INFORMATION, TABLE S2 RIGHTS AND PERMISSIONS Reprints and permissions ABOUT THIS
ARTICLE CITE THIS ARTICLE Ma, C., Sacco, M.D., Hurst, B. _et al._ Boceprevir, GC-376, and calpain inhibitors II, XII inhibit SARS-CoV-2 viral replication by targeting the viral main
protease. _Cell Res_ 30, 678–692 (2020). https://doi.org/10.1038/s41422-020-0356-z Download citation * Received: 07 May 2020 * Accepted: 29 May 2020 * Published: 15 June 2020 * Issue Date:
August 2020 * DOI: https://doi.org/10.1038/s41422-020-0356-z SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a
shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative