Role of protein kinase c and nf-κb in proteolysis-inducing factor-induced proteasome expression in c2c12 myotubes
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT Proteolysis-inducing factor (PIF) is a sulphated glycoprotein produced by cachexia-inducing tumours, which initiates muscle protein degradation through an increased expression of
the ubiquitin–proteasome proteolytic pathway. The role of kinase C (PKC) in PIF-induced proteasome expression has been studied in murine myotubes as a surrogate model of skeletal muscle.
Proteasome expression induced by PIF was attenuated by 4_α_-phorbol 12-myristate 13-acetate (100 nM) and by the PKC inhibitors Ro31-8220 (10 _μ_ M), staurosporine (300 nM), calphostin C (300
nM) and Gö 6976 (200 _μ_ M). Proteolysis-inducing factor-induced activation of PKC_α_, with translocation from the cytosol to the membrane at the same concentration as that inducing
proteasome expression, and this effect was attenuated by calphostin C. Myotubes transfected with a constitutively active PKC_α_ (_p_CO2) showed increased expression of proteasome activity,
and a longer time course, compared with their wild-type counterparts. In contrast, myotubes transfected with a dominant-negative PKC_α_ (pKS1), which showed no activation of PKC_α_ in
response to PIF, exhibited no increase in proteasome activity at any time point. Proteolysis-inducing factor-induced proteasome expression has been suggested to involve the transcription
factor nuclear factor-_κ_B (NF-_κ_B), which may be activated through PKC. Proteolysis-inducing factor induced a decrease in cytosolic I-_κ_B_α_ and an increase in nuclear binding of NF-_κ_B
in _p_CO2, but not in pKS1, and the effect in wild-type cells was attenuated by calphostin C, confirming that it was mediated through PKC. This suggests that PKC may be involved in the
phosphorylation and degradation of I-_κ_B_α_, induced by PIF, necessary for the release of NF-_κ_B from its inactive cytosolic complex. SIMILAR CONTENT BEING VIEWED BY OTHERS DUAL ROLES OF
MTORC1-DEPENDENT ACTIVATION OF THE UBIQUITIN-PROTEASOME SYSTEM IN MUSCLE PROTEOSTASIS Article Open access 27 October 2022 ACTIVATORS OF THE 26S PROTEASOME WHEN PROTEIN DEGRADATION INCREASES
Article Open access 08 January 2025 THE PHOSPHOENOLPYRUVATE CARBOXYKINASE (PEPCK) INHIBITOR, 3-MERCAPTOPICOLINIC ACID (3-MPA), INDUCES MYOGENIC DIFFERENTIATION IN C2C12 CELLS Article Open
access 17 December 2020 MAIN Loss of skeletal muscle in cancer cachexia results in asthenia, immobility and eventually death through impairment of respiratory function. Nutritional
supplementation alone is unable to reverse this wasting process (Evans et al, 1985), suggesting that the balance between protein synthesis and degradation is impaired, especially since
visceral protein reserves are preserved and may even increase (Fearon, 1992). Thus, while protein synthesis in skeletal muscle of cachectic patients is impaired (Lundholm et al, 1976), there
is also an increase in protein degradation (Lundholm et al, 1982). An increased activity of the ubiquitin–proteasome proteolytic pathway is considered to be the major factor for this
increased protein degradation (Williams et al, 1999). Tumour production of a sulphated glycoprotein called proteolysis-inducing factor (PIF) may be responsible for the progressive loss of
skeletal muscle in cancer cachexia (Todorov et al, 1996). Proteolysis-inducing factor is only produced by cachexia-inducing tumours, and when purified and administered to mice it induces a
specific loss of skeletal muscle, while visceral protein is maintained or even increased, as in cancer cachexia (Lorite et al, 1998). Using a surrogate model of skeletal muscle, PIF was
shown to inhibit protein synthesis and increase protein degradation (Smith et al, 1999). The increased protein degradation was shown to arise from an increased expression of the
ubiquitin–proteasome proteolytic pathway (Lorite et al, 2001). Protein degradation induced by PIF was accompanied by an increased release of arachidonic acid from membrane phospholipids and
its subsequent metabolism to 15-hydroxyeicosatetraenoic acid (15-HETE), which was shown to be related to protein catabolism. Both arachidonic acid and lipoxygenase metabolites have been
shown to activate protein kinase C (PKC) (Fan et al, 1990), which may play a role in PIF-induced proteasome expression. Phosphorylation of proteasome subunits may be important in the
regulation of proteasome activity, since some proteasome subunits have potential tyrosine (Tanaka et al, 1990) and serine/threonine (Heinemeyer et al, 1994) phosphorylation sites and
dephosphorylation by acid phosphatase have been shown to lower proteasome activity significantly (Mason et al, 1996). At least one subunit (S4) has several potential PKC phosphorylation
sites (Dubiel et al, 1992). Alternatively, PKC may play a role in activation of the transcription factor nuclear factor-_κ_B (NF-_κ_B), which has been shown to be involved in the production
of interleukin-8 (IL-8), IL-6, C-reactive protein and ICAM-1 in liver cells (Watchorn et al, 2001), and may also be involved in PIF-induced proteasome expression (Whitehouse and Tisdale,
2003). Protein kinase C has been suggested as being an upstream activator of the I-_κ_B kinase complex (IKK) (Lallena et al, 1999; Trushin et al, 1999; Vertegaal et al, 2000) leading to
I-_κ_B_α_ phosphorylation and ubiquitination and the subsequent processing of the 26S proteasome followed by the translocation of NF-_κ_B into the nucleus. The present study investigates the
role of PKC in PIF-induced proteasome expression and its relationship to activation of NF-_κ_B in C2C12 murine myotubes. MATERIALS AND METHODS MATERIALS Foetal calf serum (FCS), horse serum
(HS) and Dulbecco's modified Eagle's medium (DMEM) were purchased from Invitrogen (Paisley, Scotland). Mouse monoclonal antibodies to proteasome 20S _α_-subunits were from
Affiniti Research Products (Exeter, UK). Rabbit polyclonal antisera to ubiquitin conjugating enzyme (E214k) were a gift from Dr Simon Wing, McGill University (Montreal, Canada). Rabbit
polyclonal antisera to murine I-_κ_B_α_, _β_-tubulin and PKC_α_ were from Calbiochem (Herts, UK) as were PMA, Ro31-8220, calphostin C, staurosporine and Gö 6976. Rabbit polyclonal antisera
to mouse actin were from Sigma-Aldridge (Dorset, UK). Peroxidase-conjugated goat antirabbit and rabbit antimouse secondary antibodies were from Dako Ltd (Cambridge, UK). Hybond™
nitrocellulose membranes and enhanced chemiluminescence (ECL) were from Amersham Life Science Products (Bucks, UK). Electrophoretic-mobility shift (EMSA) gel shift assay kits were from
Panomics (California, USA). _Escherichia coli_ DH5_α_ cells were from Invitrogen (Paisley, Scotland). Constitutively active and mutant plasmids of PKC_α_ were a gift from Prof. Peter Parker
(Cancer Research, UK). The insert A25E PKC_α_ is constitutively active due to a deletion of amino acids 22–28 in the N-terminal region and is expressed via the _p_CO2 vector (Pears et al,
1990). Protein kinase C _α_ (T/A)3 is a dominant-negative mutant expressed in pKS1 (Bornancin and Parker, 1996). Plasmid DNA was purified using the WIZARD® PureFection purification system
(Promega, Southampton, UK) according to the manufacturer's protocol. Primers for PCR analysis were from MWG Biotech (Ebersberg, Germany). GeneJuice™ for transfection studies was
purchased from Calbiochem (Herts, UK). TRANSFORMATION OF BACTERIA _E. coli_ DH5_α_ were transformed with both constitutively active and mutant PKC_α_ using heat shock, and selected with
ampicillin (100 _μ_g ml−1). Positive clones were identified using primers with homology to bovine PKC (forward 5′-CAC CTG TGA TAT GAA CGT GC-3′ reverse 5′-GAA GTT GAA GTC CGT GAG C-3′). The
product was about 600 bp as determined on a 2% agarose gel. Plasmid DNA was extracted from positive colonies grown overnight in an LB medium containing ampicillin (100 _μ_g ml−1).
PURIFICATION OF PIF PIF was purified from solid MAC16 tumours excised from mice with a weight loss between 20 and 25% as previously described (Todorov et al, 1996; Whitehouse and Tisdale,
2003). Tumours were homogenised in 10 mM Tris-HCl, pH 8.0, containing 0.5 mM phenylmethylsulphonyl fluoride, 0.5 mM EGTA and 1 mM dithiothreitol at a concentration of 5 ml g−1 tumour. The
supernatant obtained after addition of ammonium sulphate (40% w v−1) was subjected to affinity chromatography using anti-PIF monoclonal antibody coupled to a solid matrix. The immunogenic
fractions were concentrated and used for further studies. MYOGENIC CELL CULTURE AND TRANSFECTION The C2C12 myoblast cell line was grown in DMEM supplemented with 10% FCS plus 1% penicillin
and streptomycin under an atmosphere of 10% CO2 in air. Transfection was carried out on cells at 50% confluency using GeneJuice™ transfection reagent, according to the manufacturer's
protocol and selected by resistance to ampicillin (5 g l−1). Transfected myoblasts were stimulated to differentiate by replacing the growth medium with DMEM supplemented with 2% HS, when the
cells reached confluence. Differentiation was allowed to continue for 5–9 days until myotubes were clearly visible, and used for the experiments described in results. MEASUREMENT OF
PROTEASOME ‘CHYMOTRYPSIN-LIKE ACTIVITY ‘Chymotrypsin-like’ enzyme activity was determined fluorimetrically by the method of Orino et al (1991) as previously described (Lorite et al, 2001).
Myotubes were washed with ice-cold phosphate-buffered saline (PBS) and sonicated in 20 mM Tris-HCl, pH 7.5, 2 mM ATP, 5 mM MgCl2 and 1 mM dithiothreitol at 4°C. The supernatant formed by
centrifugation at 18 000 G for 10 min was used to measure the ‘chymotrypsin-like’ enzyme activity by the release of aminomethyl coumarin (AMC) from the fluorogenic peptide succinyl-LLVY-AMC
(0.1 mM). Activity was measured in the presence and absence of the specific proteasome inhibitor lactacystin (10 _μ_ M). Only lactacystin-suppressible activity was considered to be
proteasome specific. WESTERN BLOT ANALYSIS Cytoplasmic proteins, obtained from the above assay, were also used for Western blotting, while the pellet was dissolved in sonicating buffer
containing 0.1% Nonidet P40 and used as a source of cell membranes. Both extracts were loaded at 2–5 _μ_g protein and resolved on 10% sodium dodecylsulphate: polyacrylamide gels and
transferred to Hybond™ nitrocellulose membrane. Membranes were blocked with 5% Marvel in PBS. The primary antibodies for PKC_α_, E214k and _β_-tubulin were used at a dilution of 1 : 100,
while antibodies for I-_κ_B_α_ were at 1 : 1000 and 20S proteasome _α_-subunits at 1 : 1500. The secondary antibodies were used at a dilution of 1 : 2000. Incubation was carried out for 2 h
at room temperature, and development was by ECL. ELECTROPHORESIS MOBILITY SHIFT ASSAY DNA-binding proteins were extracted from myotubes by the method of Andrews and Faller (1991), which
utilises hypotonic lysis followed by high salt extraction of nuclei. The EMSA-binding assay was carried out using a Panomics EMSA ‘gel shift’ kit according to the manufacturer's
instructions. STATISTICAL ANALYSIS Differences as means between groups was determined by one-way ANOVA followed by Tukey–Kramer multiple comparison test. RESULTS To evaluate the role of PKC
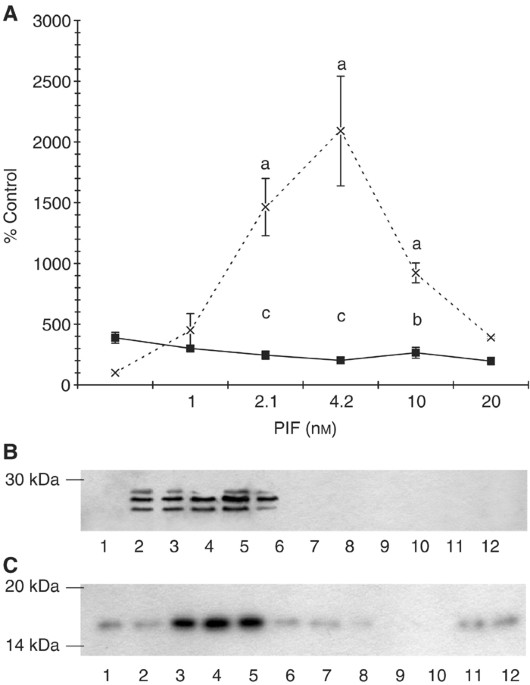
in PIF-induced proteasome expression, the effect of excess 4_α_-phorbol 12-myristate 13-acetate (PMA) on ‘chymotrypsin-like’ enzyme activity, the predominant proteolytic activity of the
proteasome (Figure 1A), and on expression of proteasome 20S_α_ subunits (Figure 1B) and the ubiquitin-conjugating enzyme (E214k) (Figure 1C) was determined in C2C12 myotubes 24 h after PIF
addition. Proteolysis-inducing factor produced an increase in ‘chymotrypsin-like’ enzyme activity, proteasome 20S_α_ subunits and E214k with a maximal effect between 2.1 and 10 nM, and this
effect was completely attenuated in myotubes pretreated with PMA. These results suggest that PKC may be important in PIF-induced proteasome expression. To confirm a role for PKC in this
process, the effect of Ro31-8220, a competitive and selective PKC inhibitor (Beltman et al, 1996), staurosporine, a broad-spectrum inhibitor of protein kinases (Couldwell et al, 1994),
calphostin C, a highly specific inhibitor of PKC (Jarvis et al, 1994), and Gö 6976, which selectively inhibits PKC_α_ and _β_, isoenzymes (Wang et al, 1998), on the PIF-induced increase in
‘chymotrypsin-like’ enzyme activity was determined (Figure 2). The PIF-induced enzyme activity was completely attenuated by 10 _μ_ M Ro31-8220 (Figure 2A), 300 nM staurosporine (Figure 2B),
300 nM calphostin C (Figure 2C) and 200 _μ_ M Gö 6976 (Figure 2D). In addition, calphostin C completely attenuated the PIF-induced increase in proteasome 20S _α_-subunit expression (Figure
3A) and E214k (Figure 3B). Proteolysis-inducing factor induced a decrease in cytosolic PKC (Figure 4A) and an increase in membrane-bound PKC_α_ (Figure 4B) at the same concentrations as
those inducing proteasome expression (Figure 1) and this effect was attenuated by both calphostin C (Figure 4A and B) and eicosapentaenoic acid (EPA) (Figure 4D). These results confirm a
role for PKC in PIF-induced proteasome expression, and suggest another mechanism by which EPA may attenuate PIF-induced protein degradation through inhibition of PKC. To further substantiate
a role for PKC in the induction of proteasome expression by PIF C2C12, myoblasts were transfected with plasmids encoding constitutively active PKC-_α_ (_p_CO2) and dominant-negative PKC-_α_
(T/A)3 (pKS1) (Bornancin and Parker, 1996; Schonwasser et al, 1998), and induced to differentiate into myotubes. Myotubes transfected with _p_CO2 showed an increased sensitivity to PIF, as
determined by the ‘chymotrypsin-like’ enzyme activity (Figure 5) in comparison with wild-type myotubes, with a significant increase within 3 h of PIF addition (Figure 5A) persisting up to 48
h (Figure 5D). In addition, the elevation of ‘chymotrypsin-like’ enzyme activity in myotubes transfected with _p_CO2 greatly exceeded that in wild type at all time points. In contrast,
myotubes transfected with the dominant-negative PKC_α_, pKS1 showed no elevation in ‘chymotrypsin-like’ enzyme activity in response to PIF at any time point (Figure 5). These results were
confirmed by Western blotting of cellular supernatants for 20S proteasome _α_-subunit expression (Figure 6A) and E214k (Figure 6B). Proteolysis-inducing factor induced an increase in both
proteasome _α_-subunit expression and E214k in _p_CO2 but not pKS1, confirming a role for PKC in this process. The ability of PIF to activate PKC_α_ in _p_CO2, but not in pKS1, was confirmed
by Western blotting (Figure 7). The concentrations of PIF causing maximum activation of PKC_α_ were the same as those inducing 20S proteasome _α_-subunit expression (Figure 6A). We have
recently shown (Whitehouse and Tisdale, 2003) that PIF-induced proteasome expression appears to require activation of NF-_κ_B. One mechanism by which PKC may function in the PIF signalling
pathway is activation of IKK with subsequent phosphorylation and degradation of I-_κ_B, and translocation of NF-_κ_B from the cytosol to the nucleus (Vertegaal et al, 2000). Evidence for
this hypothesis is provided by the following experiments. Proteolysis-inducing factor induced a decrease in cytoplasmic I-_κ_B_α_ within 30 min of addition to wild-type cells (Figure 8A),
accompanied by nuclear accumulation of NF-_κ_B (Figure 8C) and this effect was completely attenuated by calphostin C (Figure 8). In addition, PIF induced a decrease in I-_κ_B_α_ (Figure 9A)
and an increase in DNA binding of NF-_κ_B (Figure 9C) in myotubes transfected with constitutively active PKC_α_ (_p_CO2), but not in those containing dominant-negative PKC-_α_ (pKS1). These
results suggest that activation of PKC by PIF in muscle cells leads to I-_κ_B_α_ degradation, nuclear accumulation of NF-_κ_B and an increased proteasome expression leading to increased
intracellular protein degradation (Whitehouse and Tisdale, 2003). DISCUSSION Although increased intracellular protein catabolism is a common feature of many disease states, there is little
knowledge of the cellular signalling pathways involved, which may be useful in therapeutic intervention. Initial studies suggested that prostaglandin E2 (PGE2) was involved in total protein
degradation in skeletal muscle, based on the demonstration in a variety of muscle types that tyrosine release was stimulated by arachidonic acid and PGE2 (Rodemann and Goldberg, 1982) and
blocked by prostaglandin synthesis inhibitors (Strelkov et al, 1989). However, other studies (Hasselgren et al, 1990) found no evidence that total or myofibrillar protein breakdown in normal
or septic muscle is regulated by PGE2. Studies with PIF showed that total protein breakdown was related to the release of arachidonic acid and formation of PGE2, but that PGE2 was not the
eicosanoid responsible for the effect (Smith et al, 1999). Although the arachidonic acid was converted into a range of PGs and HETEs, only one metabolite 15-HETE alone was capable of
inducing protein degradation. Further studies showed that 15-HETE induced an increase in expression of the ubiquitin–proteasome pathway, which was responsible for the initiation of protein
catabolism and that this process involved the transcription factor NF-_κ_B (Whitehouse et al, 2003). The present study has investigated the possibility that PKC may act as an intermediate in
the PIF signalling pathway transmitting the rise in 15-HETE into activation of NF-_κ_B. The results support the suggestion that PKC plays a central role in the induction of proteasome
expression by PIF and thus protein degradation. Previous studies (Smith and Tisdale, 2003) have shown that PIF induces activation of phospholipase C (PLC) as an important signalling event in
inducing proteasome expression. Activation of PLC would result in the generation of diacylglycerol (DAG), which would then induce translocation of PKC from the cytosol to the membrane,
resulting in the complete activation of the kinase. Indeed, PIF has been shown to induce translocation of PKC_α_ from the cytosol to the membrane at the same concentrations as those inducing
proteasome expression. The importance of this step to the induction of proteasome expression by PIF is shown by the attenuation of this process by a range of inhibitors of PKC. In addition,
myotubes transfected with a dominant-negative mutant of PKC_α_ also showed no induction of proteasome expression in the presence of PIF. Interestingly, myotubes transfected with
constitutively active PKC_α_ showed an increased induction of proteasome expression compared with their wild-type counterparts, confirming the importance of this pathway in the signalling
cascade. At present, it is not known which particular isoenzymes of PKC are involved in this process, or indeed whether activation of PKC occurs through production of DAG via PLC or directly
through production of 15-HETE. Attenuation of PIF-induced activation of PKC provides another control point where EPA may interfere with the signalling cascade leading to increased
proteasome expression. Eicosapentaenoic acid is an effective anticachectic agent both in murine models of cachexia (Beck et al, 1991) and in weight-losing patients with pancreatic cancer
(Barber et al, 1999), and effectively attenuates PIF-induced proteasome expression in murine myotubes (Whitehouse and Tisdale, 2003). Eicosapentaenoic acid inhibits both the release of
arachidonic acid from membrane phospholipids and formation of 15-HETE in response to PIF (Smith et al, 1999), and stabilised the NF-_κ_B/I-_κ_B complex in the cytosol, preventing nuclear
accumulation of NF-_κ_B (Whitehouse and Tisdale, 2003). This study shows that EPA also attenuates PIF-induced activation of PKC, which may be due to reduced generation of DAG (Sperling et
al, 1993). In addition, DAG with an n-3 polyunsaturated fatty acid (PUFA) occupying the sn-2 position were found to be less effective in activating PKC than DAG with an n-6 PUFA, and n-3
PUFA decreased the effectiveness of activation of PKC and binding of phosphatidyl serine in the cell membrane (Terano et al, 1996). Protein kinase C _α_ is an upstream activator of the
I-_κ_B kinase complex (IKK) (Vertegaal et al, 2000), which phosphorylates I-_κ_B_α_ at serines-32 and -36 leading to ubiquitination and subsequent proteasome proteolysis. This suggests a
mechanism by which PIF may induce degradation of I-_κ_B_α_ and stimulate nuclear binding of NF-_κ_B (Whitehouse and Tisdale, 2003). Nuclear factor-_κ_B regulates the transcription of a
number of genes and has been shown (Li and Reed, 2000) to be an essential mediator of TNF-_α_-induced protein catabolism in differentiated muscle cells. This study shows that degradation of
I-_κ_B_α_ and translocation of NF-_κ_B to the nucleus in response to PIF is attenuated by calphostin C and is not seen in myotubes expressing mutant PKC_α_. This suggests that PKC acts as an
important mediator in activation of NF-_κ_B in response to PIF. It is not known whether NF-_κ_B acts alone or in concert with other transcriptional activators in PIF-induced proteasome
expression and future studies will be aimed at identifying the role of NF-_κ_B in this process. CHANGE HISTORY * _ 16 NOVEMBER 2011 This paper was modified 12 months after initial
publication to switch to Creative Commons licence terms, as noted at publication _ REFERENCES * Andrews NC, Faller DV (1991) A rapid micropreparation technique for extraction of DNA-binding
proteins from limiting numbers of mammalian cells. _Nucleic Acid Res_ 19: 2499 Article CAS Google Scholar * Barber MD, Ross JA, Voss AC, Tisdale MJ, Fearon KCH (1999) The effect of an
oral nutritional supplement enriched with fish oil on weight-loss in patients with pancreatic cancer. _Br J Cancer_ 81: 80–86 Article CAS Google Scholar * Beck SA, Smith KL, Tisdale MJ
(1991) Anticachectic and antitumor effect of eicosapentaenoic acid and its effect on protein turnover. _Cancer Res_ 51: 6089–6093 PubMed CAS Google Scholar * Beltman J, McCormick F, Cook
SJ (1996) The selective protein kinase C inhibitor, Ro 31-8220 inhibits mitogen-activated protein kinase phosphate-1 (MKP-1) expression, induces c-Jun expression and activates Jun N-terminal
kinase. _J Biol Chem_ 43: 27018–27024 Article Google Scholar * Bornancin F, Parker P (1996) Phosphorylation of threonine 638 critically controls dephosphorylation and inactivation of
protein kinase C _α_. _Curr Biol_ 6: 1114–1123 Article CAS Google Scholar * Couldwell WT, Hinton DR, He S, Chen TC, Sebat I, Weiss MH, Law RE (1994) Protein kinase C inhibitors induce
apoptosis in human malignant glioma cell lines. _FEBS Lett_ 345: 43–46 Article CAS Google Scholar * Dubiel W, Ferrell K, Pratt G, Rechsteiner M (1992) Subunit 4 of the 26S proteasome is a
member of a novel eukaryotic ATPase family. _J Biol Chem_ 267: 22699–22702 PubMed CAS Google Scholar * Evans WK, Makuch R, Clamon GH, Feld K, Weiner RS, Moran E, Blum R, Shepherd FA,
Jeejeebhoy KW, DeWys WD (1985) Limited impact of total parenteral nutrition on nutritional status during treatment for small cell lung cancer. _Cancer Res_ 45: 3347–3353 PubMed CAS Google
Scholar * Fan X, Hunag X, DaSilva C, Castagna M (1990) Arachidonic acid and related methyl ester mediate protein kinase C activation in intact platelets through the arachidonate metabolism
pathways. _Biochem Biophys Res Commun_ 109: 933–940 Article Google Scholar * Fearon KCH (1992) The mechanisms and treatment of weight loss in cancer. _Proc Nutr Soc_ 51: 251–265 Article
CAS Google Scholar * Hasselgren P-O, Zamir O, James JH, Fischer JE (1990) Prostaglandin E2 does not regulate total or myofibrillar protein breakdown in incubated skeletal muscle from
normal or septic rats. _Biochem J_ 270: 45–50 Article CAS Google Scholar * Heinemeyer W, Trondle N, Albrecht G, Wolf DH (1994) PRE5 and PRE6, the last missing genes encoding 20S
proteasome subunits from yeast? Indication for a set of 14 different subunits in the eukaryotic proteasome core. _Biochem_ 33: 12229–12237 Article CAS Google Scholar * Jarvis WD, Turner
AJ, Povrik LF, Taylor RS, Grant S (1994) Induction of apoptotic DNA fragmentation and cell death in HL-60 human promyelocitic leukaemia cells by pharmacological inhibitors of protein kinase
C. _Cancer Res_ 54: 1707–1714 PubMed CAS Google Scholar * Lallena M-J, Diaz-Meco MT, Bren G, Paya CV, Moscat J (1999) Activation of I_κ_B kinase _β_ by protein kinase C isoforms. _Mol
Cell Biol_ 19: 2180–2188 Article CAS Google Scholar * Li Y-P, Reed MB (2000) NF-_κ_B mediates the protein loss induced by TNF-_α_ in differentiated skeletal muscle myotubes. _Am J
Physiol_ 279: R1165–R1170 CAS Google Scholar * Lorite MJ, Smith HJ, Arnold JA, Morris A, Thompson MG, Tisdale MJ (2001) Activation of ATP-ubiquitin-dependent proteolysis in skeletal muscle
_in vivo_ and murine myoblasts _in vitro_ by a proteolysis-inducing factor (PIF). _Br J Cancer_ 85: 297–302 Article CAS Google Scholar * Lorite MJ, Thompson MG, Drake JL, Carling G,
Tisdale MJ (1998) Mechanism of muscle protein degradation induced by a cancer cachectic factor. _Br J Cancer_ 78: 850–856 Article CAS Google Scholar * Lundholm K, Bennegard K, Eden E,
Rennie MJ (1982) Efflux of 3-methylistidine from the leg in cancer patients who experience weight loss. _Cancer Res_ 42: 4809–4818 Google Scholar * Lundholm K, Bylund AC, Holm J, Schersten
T (1976) Skeletal muscle metabolism in patients with malignant tumour. _Eur J Cancer_ 12: 465–473 Article CAS Google Scholar * Mason GG, Hendil KB, Rivett AJ (1996) Phosphorylation of
proteasomes in mammalian cells. Identification of two phosphorylated subunits and the effect of phosphorylation on activity. _Biochemistry_ 238: 453–462 CAS Google Scholar * Orino E,
Tanaka K, Tamura T, Sone S, Ogura T, Ichihara A (1991) ATP-dependent reversible association of proteasomes with multiple protein components to form 26S complexes that degrade ubiquitinated
proteins in human HL-60 cells. _FEBS Lett_ 284: 206–210 Article CAS Google Scholar * Pears CJ, Kour G, House C, Kemp BE, Parker PJ (1990) Mutagenesis of pseudosubstrate site of protein
kinase C leads to activation. _Eur J Cancer_ 194: 89–94 CAS Google Scholar * Rodemann HP, Goldberg AL (1982) Arachidonic acid, prostaglandin E2 and F2 _α_ influence rates of protein
turnover in skeletal and cardiac muscle. _J Biol Chem_ 257: 1632–1638 PubMed CAS Google Scholar * Schonwasser DC, Marais RM, Marshall CJ, Parker PJ (1998) Activation of the
mitogen-activated protein kinase/extracellular signal-regulated kinase pathway by conventional, novel and atypical protein kinase C isotypes. _Mol Cell Biol_ 18: 790–798 Article CAS Google
Scholar * Smith HJ, Lorite MJ, Tisdale MJ (1999) Effect of a cancer cachectic factor on protein synthesis/degradation in murine C2C12 myoblasts: modulation by eicosapentaenoic acid.
_Cancer Res_ 59: 5507–5513 PubMed CAS Google Scholar * Smith HJ, Tisdale MJ (2003) Signal transduction pathways involved in proteolysis-inducing factor induced proteasome expression in
murine myotubes. _Br J Cancer_ 89: 1783–1788 Article CAS Google Scholar * Sperling RI, Benincaso AI, Knoell CT, Larkin JK, Austen KF, Robinson DR (1993) Dietary omega-3 polyunsaturated
fatty acids inhibit phosphoinositide formation and chemotaxis in neutrophils. _J Clin Invest_ 91: 651–660 Article CAS Google Scholar * Strelkov AB, Fields ALA, Baracos VE (1989) Effects
of systemic inhibition of prostaglandin production on protein metabolism in tumor-bearing rats. _Am J Physiol_ 257: C261–C269 Article CAS Google Scholar * Tanaka K, Fujiwara T, Kumatori
A, Shin S, Yoshimura T, Ichihara A, Tokunaga F, Aruga R, Iwanaga S, Kakizuka A, Nakanishi S (1990) Molecular cloning of cDNA for proteasome from rat liver: primary structure of component C3
with a possible tyrosine phosphorylation site. _Biochemistry_ 29: 3777–3785 Article CAS Google Scholar * Terano T, Shiina T, Tamura Y (1996) Eicosapentaenoic acid suppressed the
proliferation of vascular smooth muscle cells through modulation of various steps of growth signals. _Lipids_ 31: S301–S304 Article CAS Google Scholar * Todorov P, Cariuk P, McDevitt T,
Coles B, Fearon K, Tisdale M (1996) Characterization of a cancer cachectic factor. _Nature_ 379: 739–742 Article CAS Google Scholar * Trushin SA, Pennington KN, Algecirao-Schimnich A,
Paya CV (1999) Protein kinase C and calcineurin synergize to activate I_κ_B kinase and NF-_κ_B in T lymphocytes. _J Biol Chem_ 274: 22923–22931 Article CAS Google Scholar * Vertegaal ACO,
Kuiperij HB, Yamaoka S, Courtois G, van der Eb AJ, Zantema A (2000) Protein kinase C-_α_ is an upstream activator of the I_κ_B kinase complex in the TPA signal transduction pathway to
NF-_κ_B in U20S cells. _Cell Signal_ 12: 759–768 Article CAS Google Scholar * Watchorn TM, Waddell ID, Dowidar N, Ross JA (2001) Proteolysis-inducing factor regulates hepatic gene
expression via the transcription factors NF-_κ_B and STAT3. _FASEB J_ 15: 562–564 Article CAS Google Scholar * Wang D, Yu X, Brecher P (1998) Nitric oxide and _N_-acetylcysteine inhibit
the activation of mitogen-activated protein kinases by angiotensin II in rat cardiac fibroblasts. _J Biol Chem_ 273: 33027–33034 Article CAS Google Scholar * Whitehouse AS, Khal J,
Tisdale MJ (2003) Induction of protein catabolism in myotubes by 15(S)-hydroxyeicosatetraenoic acid through increased expression of the ubiquitin-proteasome pathway. _Br J Cancer_ 89:
737–745 Article CAS Google Scholar * Whitehouse AS, Tisdale MJ (2003) Increased expression of the ubiquitin–proteasome pathway in murine myotubes by proteolysis-inducing factor (PIF) is
associated with activation of the transcription factor NF-_κ_B. _Br J Cancer_ 89: 1116–1122 Article CAS Google Scholar * Williams A, Sun X, Fischer JE, Hasselgren P-O (1999) The
expression of genes in the ubiquitin–proteasome proteolytic pathway is increased in skeletal muscle from patients with cancer. _Surgery_ 126: 744–750 Article CAS Google Scholar Download
references ACKNOWLEDGEMENTS This work has been supported by the Lustgarten Foundation for Pancreatic cancer research. AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Pharmaceutical Sciences
Research Institute, Aston University, Birmingham, B4 7ET, UK H J Smith, S M Wyke & M J Tisdale Authors * H J Smith View author publications You can also search for this author inPubMed
Google Scholar * S M Wyke View author publications You can also search for this author inPubMed Google Scholar * M J Tisdale View author publications You can also search for this author
inPubMed Google Scholar CORRESPONDING AUTHOR Correspondence to M J Tisdale. RIGHTS AND PERMISSIONS From twelve months after its original publication, this work is licensed under the Creative
Commons Attribution-NonCommercial-Share Alike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/ Reprints and permissions ABOUT
THIS ARTICLE CITE THIS ARTICLE Smith, H., Wyke, S. & Tisdale, M. Role of protein kinase C and NF-_κ_B in proteolysis-inducing factor-induced proteasome expression in C2C12 myotubes. _Br
J Cancer_ 90, 1850–1857 (2004). https://doi.org/10.1038/sj.bjc.6601767 Download citation * Received: 09 December 2003 * Revised: 12 February 2004 * Accepted: 13 February 2004 * Published: 06
April 2004 * Issue Date: 04 May 2004 * DOI: https://doi.org/10.1038/sj.bjc.6601767 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get
shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative KEYWORDS *
Proteolysis-inducing factor * protein kinase C * nuclear factor-_κ_B * proteasome expression